Orrorin tugenensis
Stratigrafický výskyt: svrchní miocén, před 6 miliony let | |
|---|---|
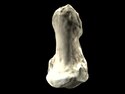
 (c) Lucius, CC BY-SA 3.0 | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | savci (Mammalia) |
| Řád | primáti (Primates) |
| Čeleď | hominidi (Hominidae) |
| Podčeleď | hominini (Homininae) |
| Tribus | Hominini? |
| Rod | Orrorin Senut et al., 2001 |
| Binomické jméno | |
| Orrorin tugenensis Senut et al., 2001 | |
| Některá data mohou pocházet z datové položky. | |
Orrorin tugenensis je druh vyhynulých hominidů, žijící ve svrchním miocénu (asi před 6,1 – 5,7 miliony let) na území dnešní Keni ve východní Africe. Vývojově stojí velmi blízko poslednímu společnému předku moderních šimpanzů a lidí a možná již patří přímo k lidské vývojové linii (subtribus Hominina). Pokud je tomu tak, je po rodu Sahelanthropus druhým nejstarším zástupce této větve. Předchází mladším homininům, jako jsou Ardipithecus a Australopithecus.
Rod Orrorin je v současnosti zastoupen jediným druhem, pojmenovaným Orrorin tugenensis. Orrorin (množné číslo Orroriek) znamená v lokálním keňském jazyce Tugen „prvotní (původní) muž“. Druhový přívlastek je odvozen z místa nálezu v pohoří Tugen Hills. První odhalené ostatky získaly přezdívku „Millenium man“ nebo „Millenium ancestor“ (česky Předek milénia), protože byl zachyceny na počátku nového tisíciletí.
Objevy



Ostatky rodu Orrorin byly objeveny teprve v roce 2000. Zachytila je francouzsko-keňská expedice Kenya Paleontology Expedition, kterou vedli paleoantropologové Martin Pickford a Brigitte Senut.[1] Místem objevu je pohoří Tugen Hills, západně od jezera Baringo na západě Keni, asi 250 km severozápadně od hlavního města Nairobi.
Všechny čtyři známé lokality – Cheboit, Aragai, Kapsomin a Kapcheberek – jsou součástí souvrství Lukeino. Drtivá většina nálezů pochází z lokality Kapsomin. Zde také Kiptalam Cheboi dne 15. října 2000 zachytil vůbec první exemplář. V následujících měsících byly objeveny další zlomky patřící nejméně pěti jedincům. Zahrnují zuby, části lebky i horních a dolních končetin.[2] S těmito nálezy se shoduje také spodní stolička, kterou Martin Pickford objevil na lokalitě Cheboit již v roce 1974.[3]
Do roku 2007 bylo identifikováno celkem 20 zlomků kostí připisovaných rodu Orrorin:[4]
|
Popis
Podle nalezených ostatků lze předpokládat, že Orrorin dosahoval výšky 115–125 cm a hmotnosti 30–50 kg. Svojí velikostí se tedy přibližně vyrovnal samici moderního šimpanze.[5]
Veškeré nálezy jsou velmi zlomkovité a poskytují poměrně málo kompletní informace. Nebyla zachycena žádná větší část tváře nebo mozkovny a také obě dostupné čelisti jsou silně poškozené. Pro srovnání se staršími i mladšími rody hominidů tak z lebky zbývají prakticky jen jednotlivé zuby.[6] Ani ty ovšem nepředstavují kompletnější vzorek chrupu jednoho nebo dvou konkrétních jedinců, ale pocházejí z různých individuí i lokalit a mírně se liší také svým stářím.[7]
Zuby
Chrup nese směs primitivních i moderních znaků, ale celkově se mnohem více podobá lidoopům než australopitékům nebo lidem.[2] Zuby (zejména stoličky) jsou opatřené silnou vrstvou skloviny. Odpovídají rostlinné stravě, nejspíše ovoci. Pouze v méně příznivých obdobích roku se Orrorin mohl uchylovat ke konzumaci tužších plodů, jako jsou semena nebo hlízy.[7]
- Řezáky jsou velké, robustní, čímž se blíží spíše lidoopům (nalezen byl pouze jeden exemplář).[1]
- Špičáky jsou menší než u většiny lidoopů, přesto zůstávají ještě poměrně dlouhé a špičaté a značně připomínají špičáky samic moderních šimpanzů.[1] Morfologie korunky pak zcela odpovídá utváření korunky u lidoopů.[2] Je na ní zachován žlábek, který je u šimpanzů a goril součástí ostřícího komplexu. Tento komplex ovšem u lidí a jejich předků (Ardipithecus, Australopithecus) spolu se zmenšením špičáků vymizel.[7]
- Třenové zuby mají dva kořeny a utváření korunky se rovněž podobá zubům goril a šimpanzů.[8]
- Stoličky jsou v poměru k velikosti těla poměrně malé, na rozdíl od všech zástupců rodu Australopithecus, kteří se vyznačovali megadoncií, tedy výrazně zvětšeným chrupem.[1] Stoličky rodu Orrorin mají nízké, zaoblené hrbolky, nízkou korunku a dlouhé kořeny. Stejně jako lidské zuby postrádají zesílený lem skloviny při dásni (cingulum), typický pro vyhynulé i žijící lidoopy.[8]
Horní končetina
Z horní končetiny byl v roce 2000 nalezen pouze jeden článek prstu a silně poškozená pažní kost.[7] Oba tyto nálezy se podobají kostem moderního šimpanze i australopitéků. Pažní kost nese výraznou hranu pro úpon vřetenního svalu (musculus brachioradialis) a článek prstu je z bočního pohledu zřetelně prohnutý.[1] Silný vřetenní sval je spojován se schopností efektivního šplhu, zatímco prohnuté články prstů umožňují pevný úchop ruky. Utváření obou kostí tudíž naznačuje, že Orrorin byl velmi dobře vybaven pro pohyb v korunách stromů podobně jako moderní lidoopi, ale také zástupci rodu Australopithecus.[3]
Dalšími výzkumy byl nalezen ještě koncový článek palce, který se podobá utváření stejné kosti u člověka.[9] Podle jeho značné délky a také podle stop po úponech svalů byl Orrorin schopný tzv. precizního gripu. To znamená, že mohl pevně spojit ukazovák a palec bříšky k sobě, což mu umožnilo obratně manipulovat i s drobnými předměty.[10] Účel této schopnosti ovšem není zcela jasný. Tradičně se předpokládá, že se vyvinula až u prvních lidí díky výrobě a používání nástrojů.[11] Není však pravděpodobné, že by Orrorin vyráběl nástroje, takže úprava jeho ruky musela mít jiný důvod. Podobně uspořádanou ruku má ale kromě lidí také pavián dželada, který díky preciznímu gripu může snadno sbírat potravu.[11] Podle Brigitte Senutové může dlouhý palec souviset i s udržováním rovnováhy a potřebou přesného úchopu při lezení po stromech.[9]
 Precizní gesto lidské ruky
Precizní gesto lidské ruky Kosti při precizním gestu
Kosti při precizním gestu Srovnání koncových článků palců různých hominidů
Srovnání koncových článků palců různých hominidů- Trojrozměrná modelace nalezeného článku palce
Dolní končetina


Z dolní končetiny rodu Orrorin je dosud známá pouze stehenní kost (femur), doložená třemi zlomkovitými exempláři.[7] Lépe zachován je ovšem jen fragment levé stehenní kosti označený kódem BAR 1002´00. Z dalších dvou nálezů nemohou být kvůli špatnému stavu dochování získány téměř žádné informace.[12]
Nález BAR 1002´00 představuje zhruba horní dvě třetiny stehenní kosti, jejíž distální konec – část při kolenním kloubu – chybí. Byl objeven rozlomený na tři kusy a částečně ohryzaný masožravou šelmou. Stejně jako další dva nalezené femury postrádá téměř celý velký chocholík (trochanter major). Přesto je možné pozorovat množství jiných znaků, z nichž Martin Pickford a Brigitte Senutová usuzují, že se Orrorin pohyboval vzpřímenou dvojnohou chůzí.[4][12]
- Hlavice (caput femoris) je téměř kulová, natočená mírně dopředu.
- Krček (collum femoris) je poměrně dlouhý a v průřezu oválný.
- Průměr hlavice je zřetelně větší než průměr krčku.
- Tuberositas glutaea (úpon pro velký sval hýžďový) je výrazný, protáhlý ve směru dlouhé osy stehenní kosti.
- Je přítomná linea intertrochanterica (uchycení pro jeden z vazů kyčelního kloubu), která je sice velmi nevýrazná, ale u lidoopů na rozdíl od lidí a australopitéků zcela chybí.
- Fossa trochanterica (jamka pro úpony svalů na vnitřní straně velkého chocholíku) je poměrně mělká.
- Na krčku je zřetelný žlábek pro musculus obturatorius externus, který je spojován s plně vzpřímeným postojem, kdy jsou nohy a trup v jedné linii. U lidoopů tento znak chybí, ale vyskytuje se u pliocenních i pleistocenních předků člověka, kteří již chodili po dvou.
Využití bipední chůze nasvědčuje i rozložení hutné kostní tkáně a trámčiny, které bylo studováno pomocí počítačové tomografie. Podobně jako u moderních lidí je u Orrorina hutná kost na spodní části krčku silnější než na jeho horní hraně.[6]
Celkově tedy Orrorin vypadal spíše jako lidoop než jako člověk, pouze stehenní kosti naznačují, že na zemi mohl chodit po dvou. Články prstů a pažní kost však zároveň ukazují, že byl ještě alespoň částečně vybavený pro obratné šplhání v korunách stromů,[1] které pro něj stále neztrácely na významu. Využíval je nejen k hledání potravy, ale také k ochraně před dravci a ke stavění úkrytů na noc.[13]
Byl vysloven názor, že s bipedií může souviset i stavba ruky umožňující jemnou manipulaci. Schopnost dvojnohé chůze mohla znamenat, že Orrorin trávil méně času v korunách stromů a častěji se pohyboval přímo po zemi. Pro šplhání, které bylo pouze občasné (například při hledání úkrytu) již nebyla tolik potřebná ruka s velmi pevným úchopem. Díky tomu se její stavba mohla přizpůsobit tak, aby více vyhovovala manipulaci s potravou a jinými předměty.[11]
Životní prostředí
Orrorin žil v blízkosti paleojezer Lukeino a Mabaget.[14] Ostatky zvířat, nalezených ve stejných sedimentech ukazují, že celou oblast pokrývala v závěru miocénu mozaikovitá krajina, v níž se střídaly otevřené travnaté plochy s malými i rozsáhlejšími úseky křovin a lesních porostů.[14] Husté, ale místy i světlejší lesy se zřejmě rozkládaly zejména v okolí řek a na březích jezer, kde byl dostatek vody.[3] Zde je také doplňovaly vlhké mokřady s bohatým podrostem.[7]
Zajímavé je, že podobné prostředí bylo rekonstruováno i na místech nálezu dalších časných homininů (Sahelanthropus, Ardipithecus).[14] Naopak se značně lišilo od otevřené savany, kterou v pliocénu a pleistocénu obývaly rody Australopithecus a Homo.[7] Uvažuje se, že právě mozaikovitá krajina mohla hrát klíčovou roli v přechodu k bipední chůzi.[14]
Stopy zubů na některých nálezech naznačují, že koncentraci koster na lokalitě Kapsomin způsobil velký masožravec, pravděpodobně kočkovitá šelma. Rovněž Orrorin se zřejmě stal její obětí.[3]
Fylogeneze
Orrorin může být jedním z nejstarších zástupců lidské vývojové linie, žijící těsně po oddělení předků člověka a šimpanze. V současnosti jej stářím předčí pouze rod Sahelanthropus z Čadu. Fylogenetická pozice rodu Orrorin ale zatím není jistá a zařazení mezi lidské předky nelze bez dalších nálezů potvrdit ani vyvrátit. Podobně jako Sahelanthropus je tudíž Orrorin často klasifikován jako blíže neurčený hominin (Hominini incertae sedis).[15][16]
Zásadním argumentem pro zařazení Orrorina mezi lidské předky je předpokládaná schopnost bipední chůze. Pro tu sice díky nálezu stehenních kostí existují pádnější doklady než u Sahelanthropa, moderněji utvářené kosti končetin ale doprovázejí zuby s mnohem starobylejším vzhledem (pravděpodobně chybí zmenšené špičáky, ale nechybí ostřící komplex). Ačkoliv primitivní znaky se ještě mohly u prvních zástupců lidské linie vyskytovat, možnost bipedie u Orrorina je sama o sobě již od oznámení nálezu v roce 2000 hojně diskutovaným až kontroverzním tématem.[17]
Námitky proti bipedii u rodu Orrorin:
- Žádný z předložených dokladů není zcela přesvědčivý, velká část důležitých znaků bipedie na nalezených kostech chybí nebo jsou málo zřetelné a navíc nekvalitně publikované.[16][18]
- Poměr velikosti hlavice a krčku stehenní kosti se blíží vyhynulým lidoopům i moderním šimpanzům, délka krčku stehenní kosti je u člověka i šimpanze téměř stejná, stejně jako úhel svíraný hlavicí (krčkem) a tělem kosti – tyto znaky proto nelze použít jako jednoznačné doklady bipedie.
- Žlábek pro musculus obturatorius externus, jeden z nejpádnějších dokladů dvojnohé chůze u rodu Orrorin se sice nevyskytuje u lidoopů, ale objevuje se u jiných primátů (pavián, chápan, hulman). Ti ovšem bipedii používají jen v malé míře a navíc se nejedná o chůzi shodnou s chůzí lidskou.[19][20]
- Studie vnitřní stavby stehenní kosti pomocí počítačové tomografie je nejednoznačná, protože snímky jsou nekvalitní, špatně čitelné a snímání bylo provedeno ze špatného úhlu.[21]
- Rozložení hutné kostní tkáně stehenní kosti není zcela shodné s lidmi ani s australopitéky (není tolik asymetrické), ale podobné schéma je možné pozorovat i u kvadrupedních primátů, například paviánů.[22]
Velká část badatelů souhlasí s tím, že Orrorin byl schopný určitého druhu dvojnohé chůze, i když není možné určit, jak často a jak efektivně ji využíval (šimpanzi také dokáží krátce chodit po dvou). Jeho způsob chůze také mohl být zcela svébytný, odlišný od člověka.[21] Téměř všichni odborníci se shodují v názoru, že v současnosti neexistuje dostatek podkladů pro určení převažujícího způsobu pohybu rodu Orrorin.[7]
Navíc i kdyby Orrorin chodil po dvou, ještě to nutně neznamená, že patřil ke skupině Hominina. Se stejným způsobem pohybu mohli v pozdním miocénu „experimentovat“ i lidoopi, kteří vyhynuli bez potomků – o bipední chůzi se uvažuje například u rodu Oreopithecus. Ani jako hominin nemusí být Orrorin předkem moderních lidí, ale pouze slepou vývojovou větví, jejíž příslušníci započatou cestu ke vzniku rodu Homo nedokončili. O tom však při současném stavu poznání nelze rozhodnout.
Kontroverzní teorii o postavení rodu Orrorin prosazují jeho objevitelé. Ti uvádějí, že stavba stehenní kosti a tudíž i způsob bipední chůze se nápadně liší od rodu Australopithecus, který je většinou paleoantropologů pokládán za přímého předka lidí. Popisují množství znaků, kterými se Orrorin mnohem víc blíží člověku.[12] Na základě tohoto pozorování předpokládají, že Orrorin je přímým předkem rodu Homo, zatímco Australopithecus byl jen slepou vývojovou větví.[3] Spojnicí mezi Orrorinem a prvními lidmi, které nyní dělí velká časová mezera zhruba 3,5 milionů let, má být dosud neobjevený rod Praeanthropus.[3] Další kandidáty na lidské předky považují za předchůdce moderních lidoopů - Ardipithecus byl součástí vývojové linie šimpanzů, zatímco Sahelanthropus patřil ke gorilám. Z dosud objevených homininů by tak mezi předky člověka patřil pouze rod Orrorin.[4]
Tuto hypotézu ovšem nepotvrdily dvě rozsáhlé srovnávací studie morfologie stehenních kostí. Ty s pomocí statistických metod hodnotily několik stovek vzorků žijících i vyhynulých hominoidů a výsledky ukazují, že femur rodu Orrorin se mnohem víc podobá kostem rodů Ardipithecus a Australopithecus než kostem člověka. To znamená, že Orrorin měl k australopitékům velmi blízko. Je možné, že patřil do skupiny Hominina a chodil po dvou. Australopithecus ale může být i nadále považován za lidského předka, protože blízký vztah rodů Orrorin a Homo nebyl prokázán.[20][23]
V každém případě jsou nalezené ostatky velmi cenným materiálem pro poznání evoluce hominidů v závěru miocénu. Významné jsou i proto, že z tohoto období, klíčového pro vznik lidské vývojové linie, existuje z Afriky dosud jen velmi málo podkladů.
Odkazy
Reference
- ↑ a b c d e f SENUT, B., PICKFORD, M., GOMMERY, D., MEIN, P., CHEBOI, K., COPPENS, Y. First hominid from the Miocene (Lukeino Formation, Kenya). Comptes Rendus de l'Académie des Sciences IIA. 2001, roč. 332, s. 137–144.
- ↑ a b c CARTMILL, M.; SMITH, F. H. The Human Lineage. New Jersey: Wiley-Blackwell, 2009. 624 s. Dostupné online.
- ↑ a b c d e f PICKFORD, M.; SENUT, B. 'Millennium ancestor', a 6-million-year-old bipedal hominid from Kenya. South African Journal of Science. 2001, roč. 97, s. 22–27.
- ↑ a b c SENUT, B.; PICKFORD, M. La dichotomie grands singes - homme revisitée. Comptes Rendus Palevol. 2004, roč. 3, s. 265–276.
- ↑ NAKATSUKASA, M., PICKFORD, M., EGI, N., SENUT, B. Femur length, body mass, and stature estimates of Orrorin tugenensis, a 6 Ma hominid from Kenya. Primates. 2007, roč. 48, s. 171–178.
- ↑ a b GALIK, K., SENUT, B., PICKFORD, M., GOMMERY, D., TREIL, J., KUPERAVAGE, A. J., ECKHARDT, R. B. External and internal morphology of the BAR 1002’00 Orrorin tugenensis femur. Science. 2004, roč. 304, s. 1450–1453.
- ↑ a b c d e f g h SAWYER, G. J.; DEAK, V. The Last Human: A Guide to Twenty-two Species of Extinct Humans. [s.l.]: Yale University Press, 2007. 256 s.
- ↑ a b SENUT, B. The Earliest Putative Hominids. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1519–1538.
- ↑ a b GOMMERY, D.; SENUT, B. La phalange distale du pouce d′Orrorin tugenensis(Miocène supérieur du Kenya). Geobios. 2006, roč. 39, s. 372–384.
- ↑ Moderní lidoopi mohou také spojit ukazovák a palec, ale ne bříšky. Mají příliš krátký a málo svalnatý palec a velmi dlouhé ostatní prsty, takže jejich manipulační schopnost nemůže být tak jemná jako u člověka.
- ↑ a b c ALMÉCIJA, S.; MOYÀ-SOLÀ, S.; ALBA, D. M. Early origin for human-like precision grasping: a comparative study of pollical distal phalanges in fossil hominins. PloS one. 2010, roč. 5, s. e11727.
- ↑ a b c PICKFORD, M. H. L., SENUT, B., GOMMERY, D., TREIL, J. Bipedalism in Orrorin tugenensis revealed by its femora. Comptes Rendus Palevol. 2002, roč. 1, s. 191–203.
- ↑ RICHMOND, B.; JUNGERS, W. Orrorin tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism. Science. 2008, roč. 319, s. 1662–1665.
- ↑ a b c d ROCHE, D., SÉGALEN, L., SENUT, B., PICKFORD, M. Stable isotope analyses of tooth enamel carbonate of large herbivores from the Tugen Hills deposits: Palaeoenvironmental context of the earliest Kenyan hominids. Earth and Planetary Science Letters. 2013, roč. 381, s. 39–51.
- ↑ WOOD, B.; HARRISON, T. The evolutionary context of the first hominins. Nature. 2011, roč. 470, s. 347–352.
- ↑ a b SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
- ↑ GIBBONS, A. Oldest human femur wades into controversy. Science. 2004, roč. 305, s. 1885.
- ↑ GIBBONS, A. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s.
- ↑ HARCOURT-SMITH, W. E. The Origins of Bipedal Locomotion. In: HENKE, W. C.; TATTERSALL, I. Handbook of palaeoanthropology 3. Berlín: Springer, 2007. S. 1483–1518.
- ↑ a b RICHMOND, B. G.; JUNGERS, W. L. Hominin proximal femur morphology from the Tugen Hills to Flores. In: REYNOLDS, S. C.; GALLAGHER, A. African Genesis: Perspectives on Hominin Evolution. Cambridge: Cambridge University Press, 2012. S. 248–267.
- ↑ a b WHITE, T. Early hominid femora: The inside story. Comptes Rendus Palevol. 2006, roč. 5, s. 99–108.
- ↑ BEGUN, D. R. The earliest hominins - is less more?. Science. 2004, roč. 303, s. 1478–1480.
- ↑ ALMÉCIJA, S., TALLMAN, M., ALBA, D. M., PINA, M., MOYÀ-SOLÀ, S., JUNGERS, W. L. The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nature communications. 2013, roč. 4.
Literatura
- GIBBONSOVÁ, A. První lidé: závody v hledání nejstaršího předka. Praha: Academia, 2011. 307 s.
- ROBERTSOVÁ, A. Evoluce: Příběh člověka. Praha: Knižní klub, 2012. 256 s.
- SVOBODA, J. Čas lovců: Aktualizované dějiny paleolitu. Brno: Akademické nakladatelství CERM, 2009. 298 s.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Orrorin tugenensis na Wikimedia Commons
Obrázky, zvuky či videa k tématu Orrorin tugenensis na Wikimedia Commons - Orrorin na webu Smithsonian Institution
Média použitá na této stránce

Autor:
- Information-silk.png: Mark James
- derivative work: KSiOM(Talk)
{kind=link}
A tiny blue 'i' information icon converted from the Silk icon set at famfamfam.com
Head of femur

Autor: Almécija, Moyà-Solà & Alba, Licence: CC BY 3.0
Morphological comparisons of pollical distal phalanges in African apes, extant humans and selected hominins.
Specimens are showed in palmar (top), oblique proximopalmar (middle) and lateral (bottom) views, and scaled to the same length to easily visualize the morphological differences. The main features related to human-like precision grasping are indicated in the middle row, whereas the palmarly protruding insertion for the flexor pollicis longus has been further signaled in lateral view (red arrows in the lower row). Note that, although with several morphological differences, all the features related to refined manipulation in modern humans are already present in the late Miocene Orrorin. By the way, the OH 7 specimen, besides its odd overall proportions, neither shows a distinctive insertion for the flexor muscle, nor a compartmentalized digital pulp. All the phalanges belong to a right thumb. Scale bars represent 5 mm.
Autor: Almécija S, Moyà-Solà S, Alba D, Licence: CC BY 2.5
360° video render of Orrorin tugenensis pollical distal phalanx.
Autor:
- Neznámý (original)
- Floris V na projektu Wikipedie v jazyce nizozemština (this version)
Location of a fossil of Australopithecus anamensis in Kenya.

Frontal longitudinal midsection of upper femur.
Autor: Gerbil, Licence: CC BY 3.0
Martin Pickford, paleontologist and paleoanthropologist

Autor: Almécija, Moyà-Solà & Alba, Licence: CC BY 3.0
Modern human thumb and index finger (right hand) during pad-to-pad precision grasping in ulnar view. The box shows the anatomy of the pollical distal phalanx and its relationship with soft structures related to refined manipulation: a huge proximopalmar fossa (orange), associated with a palmarly protruding ridge (red) for insertion of the flexor pollicis longus; a compartmentalized digital pulp to accommodate the shape of the object being manipulated; this is reflected in the presence of an ungual fossa (green), associated to the large and mobile proximal pulp, as well as a wide apical tuberosity associated with the smaller and less mobile distal pulp; and finally, the ungual spines (yellow), where the collateral intraosseous ligaments that sustain the nail bed insert.
(c) Lucius, CC BY-SA 3.0
Orrorin tugenensis fossils.
- A: BAR 1002'00, left femur;
- B: BAR 1215'00, fragmentary right proximal femur;
- C: BAR 1003'00, proximal left femur;
- D: BAR 1004'00, right distal humerus;
- E: Teeth;
- F: BAR 1000'00, left mandibular fragment with M2–3;
- G: BAR 1000'00, right mandibular fragment with M3.
Autor: Chartep, Licence: CC BY-SA 3.0
Orrorin, paleoanthropological localities, Tugen Hills, Kenya