Ploštěnci
Stratigrafický výskyt: spodní paleozoikum – současnost | |
|---|---|
 Rozmanitost ploštěnců (odshora ve směru hodinových ručiček): jednorodý diplozoid Eudiplozoon nipponicum, tasemnice dlouhočlenná (Taenia solium), motolice jaterní (Fasciola hepatica), ploštěnka Pseudobiceros hancockanus | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Podříše | Eumetazoa |
| (nezařazeno) | prvoústí (Protostomia) |
| Kmen | ploštěnci (Platyhelminthes) Gegenbaur, 1859 |
| Podkmeny | |
| |
| Některá data mohou pocházet z datové položky. | |
Ploštěnci (Platyhelminthes) jsou kmen prvoústých živočichů. V rámci tradičního systému zahrnují třídu volně žijících ploštěnek (Turbellaria) a parazitické třídy motolice (Trematoda), tasemnice (Cestoda) a jednorodí (Monogenea). Moderní systematika dělí ploštěnce na klady Catenulida (řetěznatky) a Rhabditophora, protože podle molekulárních dat parazitické třídy vytvářejí monofyletickou skupinu Neodermata, jež je vnořena v rámci původních ploštěnek. Ploštěnci se objevili zřejmě již ve spodním paleozoiku a dnes jde o velmi úspěšnou, kosmopolitně rozšířenou skupinu, jež zahrnuje přes 26 000 popsaných druhů.
Ploštěnci jsou podlouhlí, dvoustranně souměrní. Jejich tělo vyplňuje pouze řídká tkáň mezenchym, přičemž velké mezibuněčné prostory tvoří komplex tekutinou naplněných štěrbinek a dutinek. Chybí oběhová a dýchací soustava. Transport látek zajišťuje difuze, pro její usnadnění mají ploštěnci obecně zploštěné tělo a často bohatě větvený systém střev. Trávicí soustava je neúplná, bez řitního otvoru. Vylučovacími orgány jsou protonefridia, jejichž funkce je původně osmoregulační. Pokožku ploštěnek tvoří bičíkatá epitelová tkáň s různě rozmístěnými žlázovými buňkami a nervovými zakončeními. U parazitických tříd však tělo kryje neodermis, jedinečný syncytiální pokožkový kryt. Nervová soustava ploštěnců vykazuje koncentraci nervových uzlin do hlavové části těla.
Ploštěnky se většinou živí dravě anebo mrchožravě, potravu pohlcují pomocí ústního otvoru, na který navazuje svalnatý, mnohdy vychlípitelný hltan. Větší kořist mohou ploštěnky přemoci pomocí lepivých sekretů nebo jedu. Jednorodí jsou primárně vnější parazité a živí se buď pokožkou, nebo krví svých hostitelů. Motolice a tasemnice jsou parazity vnitřních orgánů, z toho motolice konzumují tkáně a tělní tekutiny hostitele, tasemnice přijímají živiny jen prostřednictvím neodermis.
Ploštěnci jsou převážně hermafrodité, ale některé ploštěnky jsou gonochoristé a z parazitických skupin představují gonochoristy také motolice krevničky. Ploštěnci se rozmnožují vajíčky. Reprodukční biologie je však velmi komplikovaná. Při pohlavním rozmnožování ploštěnek většinou dochází ke vzájemnému oplodnění dvou jedinců, jindy může oplodnění probíhat pouze jednosměrně. Možností je i samooplození nebo nepohlavní rozmnožování, například pomocí fragmentace samotného těla; ploštěnky jsou obecně známé velkou schopností regenerace. Ještě složitější je rozmnožování u parazitických tříd. Zvláště vnitřní parazité demonstrují složité vývojové cykly zahrnující několik hostitelů, střídání nepohlavního a pohlavního rozmnožování a několika tělních plánů v různých fázích životního cyklu. S hostiteli navíc parazitičtí ploštěnci často cíleně manipulují a poskytují mnoho klasických ukázek rozšířeného fenotypu.
Systematika
Vnitřní systematika
Ploštěnce (Platyhelminthes) v rámci zoologické nomenklatury poprvé klasifikoval již švédský přírodovědec Carl Linné, jenž je ve svém prvním vydání díla Systema naturae z roku 1735 považoval za součást širšího taxonu Vermes (červi). Vermes zahrnovali velké množství vzájemně nepříbuzných bezobratlých, a šlo tedy o nepřirozený, polyfyletický taxon. Německý přírodovědec Carl Vogt roku 1851 rozčlenil taxon Vermes na několik samostatných skupin: Annelida, Rotatoria, Nematelmia a Platyelmia. Taxon Platyelmia zahrnoval současné ploštěnce a vyjma nich i pásnice (Nemertea). Roku 1859 jiný německý přírodovědec, Carl Gegenbaur, použil pro taxon Platyelmia pojmenování Platyelminthes, jež je v současnosti, s přepisem Platyhelminthes, považováno za platné. Spolu s tím však Gegenbaur rovněž znovu vytyčil nepřirozený taxon Vermes, ploštěnci byli tedy na samostatný kmen povýšeni teprve později. Od roku 1876 se mezi ploštěnce neřadí pásnice, jež byly vyděleny americkým biologem Charlesem Minotem.[1][2][3]
Zmínku si zaslouží rovněž postavení některých samostatných skupin, ještě ke konci 20. století považovaných za zástupce ploštěnců. Jedná se především o skupinu praploštěnců (Acoelomorpha), primitivních mořských bezobratlých, již tvoří bezstřevky (Acoela) a praploštěnky (Nemertodermatida). Praploštěnci byli od ploštěnců vyděleni teprve na přelomu konce 20. a začátku 21. století.[4] Mezi ploštěnce byli původně řazeni i mlžojedi (Xenoturbellida), jejichž první zástupce byl popsán roku 1949 jakožto primitivní ploštěnka, resp. bazální ploštěnec.[5][6] Praploštěnci a mlžojedi dnes nejsou považováni za blízce příbuzné ploštěnců, sdružováni jsou do společného kmene Xenacoelomorpha buďto na bází bilaterií,[7] anebo v rámci druhoústých živočichů.[8][5]
Při vynechání těchto skupin tradiční systematika uznávala tři, případně čtyři třídy ploštěnců: ploštěnky (Turbellaria), motolice (Trematoda), tasemnice (Cestoda) a případně i jednorodé (Monogenea), známé i pod jménem žábrohlísti. Tento tradiční systém, ačkoli se s ním lze setkat i v modernější odborné literatuře, není přirozený. Podle molekulárních analýz vycházejí tradiční ploštěnky jako parafyletický taxon, nezahrnující tedy všechny potomky společného předka skupiny. Tradiční třídy parazitických ploštěnců jsou v monofyletickém pojetí systému vnořeny v rámci původních Turbellaria. Moderní systematika uznává dva velké klady ploštěnců, Catenulida a Rhabditophora. Klad Rhabditophora je početnější a zahrnuje i všechny notoricky známé parazitické druhy, které společně vytvářejí klad Neodermata.[9] Klady Catenulida a Rhabditophora jsou samy o sobě morfologicky dobře definované, avšak nesdílejí žádné známé morfologické apomorfie. Molekulární analýzy nicméně ploštěnce sdružující skupiny Catenulida a Rhabditophora prakticky jednoznačně považují za monofyletické.[10]
Podle molekulárních analýz vychází jakožto bazální skupina rhabditofor klad „ploštěnek“ Macrostomorpha. Vůči kladu Neodermata pak jako sesterská skupina vystupuje rod sladkovodních „ploštěnek“ Bothrioplana, resp. řád Bothrioplanida. Diskutována je příbuznost jednotlivých parazitických tříd v rámci kladu Neodermata. Tasemnice mohou tvořit sesterský taxon jak k motolicím, tak k jednorodým. Pokud tvoří tasemnice a jednorodí sesterské skupiny, pak tento klad nese pojmenování Cercomeromorpha.[10] Některé studie, jako například ty rekonstruující evoluční historii neodermátů nebo analyzující transkriptomová data, však tuto systematiku neuznávají a jednorodé naopak považují za bazální neodermáty. Skupina jednorodých může být zároveň nejspíše parafyletická.[11][12]
Následující vnitřní systematika ploštěnců a přidružený kladogram vychází z [13] a [10]:
kmen ploštěnci (Platyhelminthes)
- podkmen Catenulidea
- třída řetěznatky (Catenulida)
- podkmen Rhabditophora
- infrakmen Macrostomorpha
- řád Haplopharyngida
- řád velkoústí (Macrostomida)
- infrakmen Trepaxonemata
- nadtřída Amplimatricata
- řád mnohovětevní (Polycladida)
- řád Prorhynchida
- nadtřída Gnosonesimora
- řád Gnosonesimida
- nadtřída Euneoophora
- třída rovnostřevní (Rhabdocoela)
- řád Dalytyphloplanida
- řád Kalyptorhynchia
- třída nevětvení (Proseriata)
- řád Unguiphora
- řád Lithophora
- třída Acentrosomata
- podtřída Adiaphanida
- řád trojvětevní (Tricladida)
- řád Prolecithophora
- řád Fecampiida
- podtřída Bothrioneodermata
- infratřída Bothrioplanata
- řád Bothrioplanida
- infratřída Neodermata
- infratřída Bothrioplanata
- podtřída Adiaphanida
- třída rovnostřevní (Rhabdocoela)
- nadtřída Amplimatricata
- infrakmen Macrostomorpha
Vnější systematika
Ploštěnci byli z hlediska vnější systematiky v průběhu historie sdružováni s rozličnými skupinami živočichů, případně byli umisťováni jako bazální zástupci bilaterií. V současnosti bývají považováni za sesterskou skupinu břichobrvek (Gastrotricha) jako zástupci kladu Rouphozoa.[pozn. 1] Lze se setkat se systematikou, kdy jsou ploštěnci a břichobrvky řazeni jako zástupci širšího kladu Platyzoa, jenž zahrnoval mj. i vířníky (Rotifera), vrtejše (Acanthocephala)[pozn. 2] a čelistovky (Gnathostomulida). Tento koncept představil Thomas Cavalier-Smith roku 1998, nicméně pravděpodobně je parafyletický.[14] Kladogram znázorňující možné příbuzenské vztahy v rámci hlavních skupin spiralií je uveden níže. Sesterskou skupinou k Rouphozoa zde vychází klad Lophotrochozoa, zahrnující z významných kmenů např. měkkýše (Mollusca) a kroužkovce (Annelida).[16] Některé novější studie (2019) naznačují příbuznost ploštěnců s kroužkovci a pásnicemi v rámci lophotrochozoí.[17]
| Spiralia |
| ||||||||||||||||||||||||
Evoluce

Původ a evoluční historie ploštěnců jsou z velké části hádankou. Problém je jejich extrémní anatomická jednoduchost, kvůli níž nelze provést spolehlivou fylogenetickou analýzu založenou na morfologii, souběžně s tím nedostatek jednoznačných synapomorfií, a velmi chudé jsou rovněž samotné fosilní nálezy ploštěnců, z nichž by tyto analýzy měly vycházet. Ač se téměř jednoznačně uznává, že původní ploštěnci byli pravděpodobně volně žijící (a neparazitičtí), většinu fosilií zahrnují nepřímé důkazy o přítomnosti parazitických zástupců neodermátů v různých hostitelích.[18][19]
Tělní stavbu hypotetických předků moderních ploštěnců však lze odhadnout na základě morfologie současných bazálních linií Catenulida (řetěznatky) a Macrostomorpha. Předky dnešních ploštěnců byly nejspíše ploštěnky s jednoduchým hltanem a vakovitými střevy bez divertiklů, jednovrstevnou, obrvenou epidermis, endolecitálními vajíčky (tj. primitivnější způsob zásobování vajíček, která obsahují velké množství žloutku) a spirálním rýhováním zygoty. Ploštěnci prakticky zcela postrádají tělní dutinu, pozorovanou u ostatních trojlistých živočichů. Podle některých starších hypotéz by mohli představovat příklad pedomorfních živočichů, vycházejících z předků s pravou tělní dutinou. Pokud by tyto starší pohledy byly pravdivé, ploštěnci by mohli pocházet například z raných vývojových stádií některých prvoústých, u nichž došlo k rychlé pohlavní zralosti u larválních nebo jiných stádií, a to ještě před vznikem pravé tělní dutiny v průběhu embryonálního vývoje.[18][19]
Další otázku představuje přechod ploštěnců k parazitickému způsobu života. Společným předkem kladu Neodermata mohla být volně žijící „ploštěnka“, zřejmě všežravá. Mohla představovat mrchožrouta, anebo mikropredátora. Podle studie publikované v roce 2010 mohli prvními parazity mezi ploštěnci být někteří zástupci jednorodých (žábrohlístů), kteří se ještě živili epitelem rybích hostitelů. Postupně došlo k vyvinutí příchytných orgánů, jakož i k přechodu k potravě založené na krvi hostitele. Společný předek motolic a tasemnic byl již pravděpodobně žábrohlíst živící se krví; obě skupiny následně prodělaly nezávislý vývoj, během něhož došlo k rozvinutí odlišných potravních a rozmnožovacích strategií. Podobný pohled zároveň vylučuje postavení tasemnic a žábrohlístů jako sesterských skupin.[11]
Nejstarší fosilní záznamy hodnocené jako pozůstatek některých ploštěnců byly datovány již do spodního paleozoika,[19] z toho možné pozůstatky ploštěnek byly identifikovány ve svrchněordovických nalezištích na ostrově Anticosti.[20] Ze svrchního siluru pocházejí nálezy měkkýšů, jejichž fosilie vypadají, jako by byly během života napadeny gymnofalidními motolicemi. Tyto závěry však nejsou věrohodné, protože jedním z hostitelů gymnofalidních motolic jsou bahňáci, přičemž během siluru se ještě neobjevují ani zástupci tetrapodů. Spolehlivější rané nepřímé důkazy, potvrzující existenci ploštěnců, resp. jejich parazitických linií, pocházejí z devonu: jde o nálezy háčků na fosiliích trnoploutvých a pancířnatců, které pravděpodobně náležely jednorodým.[18][21] Jednoznačně prokazatelným nálezem je objev vajíček tasemnic v koprolitu žraloka ze svrchního permu, jenž dokazuje, že již na konci paleozoika žili ploštěnci se složitými životními cykly, kteří byli v parazitickém vztahu s tehdejšími obratlovci.[22] Nálezy ploštěnců parazitujících na suchozemských obratlovcích pak pocházejí ze spodní křídy (koprolity archosaurů).[21]
Charakteristika

Ploštěnci představují podlouhlé bilaterální živočichy s dobře vyvinutou hlavovou částí (přídí). Projevuje se u nich vývoj třetího zárodečného listu mezodermu, chybí však coelom.[23] Prostor mezi epidermis ektodermálního původu a střevy entodermálního původu je obvykle vyplněn mezenchymem, jednoduchou řídkou tkání mající charakter parenchymu, v níž jsou uloženy vnitřní orgány. Velké mezibuněčné prostory pak tvoří komplex tekutinou naplněných štěrbinek a dutinek, podílejících se díky principu nestlačitelnosti kapalin na vytvoření hydrostatického skeletu. Pod pokožkou vzniká kožně-svalový vak mezodermálního původu, který tvoří svrchní vrstva okružní svaloviny, pod níž je uložena vrstva svalů podélných, případně jsou doplněny i svaly šikmé.[24][25] U některých druhů lze v tělní stěně pozorovat i některé vápenaté struktury.[26]


Ploštěncům schází oběhová i dýchací soustava. Dýchají celým povrchem těla, transport látek zajišťuje difuze v mezenchymu.[24] Endoparazitičtí ploštěnci jsou většinou schopni jak aerobního, tak anaerobního metabolismu, což jim umožňuje přežívat v částech těl hostitelů s nedostatkem kyslíku. Konečnými produkty metabolismu v takovém případě bývají některé redukované produkty, jako je laktát nebo sukcinát.[27] Ploštěnci potřebují zachovávat ideální poměr povrchu a objemu těla, jenž umožňuje udržovat většinu životních funkcí, což dává těmto tvorům jejich charakteristický plochý tvar. Především menší druhy mohou mít tělo i válcovité, nicméně u větších zástupců – především pak u endoparazitů – se tělo postupně dorzoventrálně zplošťuje a může mít až stužkovitý tvar, což usnadňuje výměnu plynů přes tělní stěnu mezi tkáněmi a okolním prostředím.[28] Ploštěnky dosahují velikosti 1 mm až asi 30 cm,[29] zatímco tasemnice, nejdelší známí ploštěnci (a zároveň endoparazité), mohou dosahovat délky i přes 27 m.[30]
Tělo ploštěnců je u většiny druhů nesegmentované. Specializovanou stavbu těla lze pozorovat u tasemnic, které mají v případě většiny druhů skolex (hlavičku) s příchytnými orgány, krátký krček a strobilu, samotné „tělo“ tasemnice, jež je tvořeno jednotlivými články nazývanými proglotidy. Proglotid může strobila nést několik jednotek či desítek, u některých druhů, jako je tasemnice bezbranná (Taenia saginata), však i několik tisíc. Proglotidy vznikají během celého života tasemnice v těle definitivního hostitele. Na rozdíl od segmentace většiny jiných živočichů však nedorůstají z distální strany, ale od oblasti krčku, jenž obsahuje množství kmenových buněk, které proliferací dávají vzniknout novým proglotidám.[28][31] Ne všechny tasemnice však mají typicky článkované tělo, zcela nečlánkovanými zástupci jsou například jeseterovky (Amphilinidea).[32]
Krycí soustava a pohyb
.jpg)
Pokožka (epidermis) volně žijících ploštěnců je tvořena celkově nebo částečně bičíkatým epitelem s různě rozmístěnými žlázovými buňkami ektodermálního původu a nervovými zakončeními. Řetěznatky (Catenulida) mají buňky jenom s malým množstvím bičíků, což je považováno za původní znak, ačkoli buňky jejich hltanu už mají běžné obrvení.[33][34] Bičíky pomáhají přihánět k povrchu těla čerstvou okysličenou vodu[25] a u většiny benticky žijících ploštěnců činnost řasinek umožňuje také pohyb živočicha, ačkoli některé ploštěnky se při pohybu spoléhají i na svalové kontrakce, a mohou dokonce i krátce plavat.[26]
Žlázové buňky ploštěnců jsou často uloženy v mezenchymu, ústí nicméně k povrchu těla. Jejich význam spočívá v produkci sekretů, jež slouží k mnohým účelům. Mnoho druhů bentických ploštěnek má zvýšený počet těchto buněk na spodní straně těla. Buňky v takovém případě vylučují sliz, který napomáhá v pohybu po površích. Některé sekrety naproti tomu usnadňují zachycení a pozření kořisti, jiné vytvářejí kolem těla vlhký obal, který může obojživelným ploštěnkám sloužit jako ochrana před vyschnutím a zároveň napomáhat difuzi plynů. Jiné komplexy žlázových buněk umožňují přilnutí k povrchům a zahrnují proto buňky, které produkují granule s adhezivními složkami, stejně jako granule, které pomáhají toto spojení zase přerušit.[35][33] Celý proces může být velmi rychlý; například ploštěnka Macrostomum lignano má na spodní straně těla asi 130 adhezivních orgánů, které tvoří celkem tři typy buněk s různými funkcemi. Jejich součinností může ploštěnka přilnout k podkladu a zase se od něj odpoutat i několikrát za sekundu.[36] Ploštěnky podkmene Rhabditophora jsou vyjma sekretů žlázových buněk schopny vytvářet i specializované tyčinkovité inkluze nazývané rhabdoidy a rhabdity, které vznikají v mezenchymu, anebo v pokožce; po vyloučení se z nich stává slizovitá vrstva, jež může sloužit ke znehybnění kořisti, ochraně před vyschnutím, anebo jako obrana proti predátorům.[37] Několik desítek ploštěnek, například určití zástupci rovnostřevných a mnohovětevných, mohou od své kořisti získávat žahavé buňky (nematocyty), které pak využívají k obraně. Některé mnohovětevné ploštěnky jsou schopny sekretovat kyselinu.[33]

Jedinečný pokryv těla, jenž jim vynesl i jméno, mají zástupci parazitického kladu Neodermata. Zatímco jejich larvální stádia mají klasickou epidermis, alespoň částečně obrvenou, později ji u nich nahrazuje pokožkový kryt nového typu. Ten je nazýván neodermis nebo tegument a má syncytiální charakter. Ačkoli je klad/infratřída Neodermata řazena v rámci podkmene Rhabditophora, přítomnost rhabditů u neodermátů nebyla spolehlivě prokázána a v průběhu evoluce specifického tělního pokryvu byl jejich rozvoj zřejmě potlačen. Tegument představuje novou, důležitou adaptaci, která parazitům poskytuje ochranu, společně s tím usnadňuje výměnu plynů a vylučování odpadních látek difuzí a napomáhá vstřebávání živin (např. pomocí pinocytózy). Tasemnice živiny dokonce přijímají výhradně prostřednictvím neodermis.[38][35] Vzhledem k absenci obrvení pokožky u dospělců – larvy mohou být naopak vysoce pohyblivé – se parazitičtí zástupci, jako například motolice, musí při pohybu spoléhat na svalové kontrakce, endoparazité mohou být rovněž přenášeni pomocí tělních tekutin hostitele. Přichycení na hostiteli či v něm napomáhají různé příchytné struktury: skolex u tasemnic, takzvaný opisthaptor u žábrohlístů (ten je zachován po celý život[39]) či přísavky u dvojrodých motolic.[26]
Vylučovací soustava

Ploštěnci již mají specializovanou vylučovací soustavu, přičemž vylučovacími orgány jsou protonefridia, která jsou spojena sběrnými kanálky, jež ústí skrze jeden nebo více otvorů na povrchu těla (nefridiopóry).[27] Zvláště u motolic lze v rámci vylučovací soustavy pozorovat akumulace, kdy dochází ke zmenšování počtu kanálků, ale především ke vzniku jediného vylučovacího otvoru, jemuž předchází primitivní sběrací měchýřek.[40] Sběrný měchýřek se objevuje i v poslední proglotidě mladých tasemnic, ale jen do doby odvržení tohoto článku.[27]
Protonefridia se skládají z plaménkových buněk (cyrtocytů), na které se napojují buňky kanálkové.[24] Komplex charakterizuje velká vnitřní dutina (ampula), jež uvnitř obsahuje bičíky, přičemž jejich počet se může různit. Stěny jsou perforované podélnými štěrbinami, vzniklými vzájemným prostoupením buněčných výrůstků cyrtocytů a první kanálkové buňky. Kmit bičíků („plaménku“) žene tekutinu do odvodného kanálku, čímž se zároveň vytváří podtlak uvnitř ampuly, pomocí něhož dochází k nasávání další tekutiny.[25] Vnitřní obrvení mohou mít někdy i vývodné kanálky.[24]
Funkce protonefridií je původně osmoregulační, jakožto snaha vyrovnat se s hypotonickým prostředím, protože někteří starobylí ploštěnci, obývající více méně izotonické prostředí, protonefridia nepotřebují. Vylučování zplodin metabolismu představuje teprve druhotnou funkci protonefridií,[25] většina metabolických odpadů je jinak vylučována přes tělesnou stěnu.[27]
Trávicí soustava a přijímání potravy
_anterior.jpg)
Trávicí soustava ploštěnců je neúplná (tj. bez řitního otvoru), potravu ploštěnci většinou přijímají a nestrávené zbytky vyvrhují jedním otvorem.[25] Hlavní výjimku v tomto případě představují tasemnice, protože trávicí soustavu zcela redukovaly a živiny vstřebávají pouze prostřednictvím tegumentu, čemuž napomáhá systém mikroklků, jež zvětšují povrch těla pro absorpci živin.[41] Poloha ústního otvoru je v rámci ostatních ploštěnců variabilní, může ležet z ventrální i terminální strany a jak v přední části těla zvířete, tak v části zadní. Za ústním otvorem následuje svalnatý hltan (pharynx). Ten je ektodermálního původu a může obsahovat některé žlázy, které sekretují hlen, případně i některé trávicí enzymy. Hltan se u ploštěnců vyvíjí v několika typech. Nejjednodušší typ hltanu, pharynx simplex, je rourovitý a v podstatě pouze spojuje ústní otvor se střevy. Objevuje se především u ploštěnek živících se různými mikroorganismy (například řetěznatky). Složitějším typem hltanu je hltan skládaný, pharynx plicatus. Stěny takového hltanu jsou zřasené a hltan je vychlípitelný, na základě čehož je nejčastější adaptací dravých zástupců. Třetím nejčastějším typem hltanu je hltan svalnatý, pharynx bulbosus, což je silně osvalený vychlípitelný hltan ve tvaru cibule či hlízy, jenž se objevuje převážně u dravých a parazitických druhů.[42][43]

Střeva jsou mnohdy bohatě větvená, aby mohla rozvádět živiny do celého těla namísto oběhové soustavy (tzv. gastrovaskulární soustava). Způsob větvení střev některým skupinám ploštěnců vynesl i jméno. U ploštěnek mnohovětevných (Polycladida) se střeva začínají větvit za hltanem, přičemž postupným stupňovitým větvením vzniká bohatý střevní labyrint. Ploštěnky trojvětevné (Tricladida) mají střeva dokonce o třech hlavních větvích, které se následně druhotně rozvětvují skrze celé tělo.[44] Trávení ploštěnců probíhá často extracelulárně, uvnitř střeva, dokud není částečně strávený materiál fagocytován jedinou vrstvou střevní buněčné výstelky, pak již trávení pokračuje intracelulárně. U některých zástupců se objevuje i vnější trávení za pomocí enzymů vylučovaných svalnatým hltanem.[42]

Ploštěnky jsou většinou dravé, anebo mrchožravé. Většina druhů požírá malou potravu, jako jsou prvoci, malí měkkýši, korýši nebo červi. Některé ploštěnky jsou výhradními konzumenty pouze určité potravy, například houbovců, pláštěnců, svijonožců nebo mechovců. Suchozemské druhy mohou lovit žížaly nebo některé plže.[45] Velkou a pohyblivou kořist mohou ploštěnky znehybnit a pozřít mnohými způsoby. Některé produkují lepivý hlen, jenž může mít i narkotické či toxické účinky. Výzkum jedné mnohovětevné ploštěnky z Guamu ukázal, že ploštěnka k polapení pohyblivé kořisti využívá tetrodotoxin (TTX), jed známý mimo jiné od čtverzubců rodu Takifugu nebo chobotnic rodu Hapalochlaena. Několik druhů ploštěnek používá ostrý kopulační orgán k probodnutí kořisti, případně i k injikování jedu.[46][47] Některé ploštěnky žijí komenzálně až poloparaziticky, jako zástupci cizopasek (Temnocephalida), již se pomocí břišní přísavky a ouškovitých výběžků hlavové části uchycují na různých hostitelích.[48] Jiné ploštěnky dokonce žijí v symbiotickém vztahu se řasami, kdy řasám poskytují dusíkaté látky a ochranu, zatímco fotobiont produkuje sacharidy a tuky.[45] Podobné vztahy lze pozorovat například mezi zelenými řasami rodu Chlorella a sladkovodními ploštěnkami z rodu Dalyellia. Fotosymbióza se řasami je mezi trojlistými živočichy obecně vzácný fenomén, objevuje se prakticky pouze u ploštěnců a některých měkkýšů.[49] U rovnostřevných ploštěnek Baicalellia solaris a Pogaina paranygulgus se (pravděpodobně nezávisle) vyvinula kleptoplastie: z pohlcených rozsivek různých druhů si ponechávají jejich plastidy jako vlastní vnitrobuněčné organely – kleptoplastidy, aniž by došlo k jejich intracelulárnímu strávení; funkční fotosyntéza v nich přetrvává přibližně týden.[50] Řetěznatky Paracatenula spp. dokonce zcela postrádají ústní otvor, hltan a střeva. Namísto toho jejich tělo zahrnuje specializovanou tkáň zvanou trofosom, jež je složena z bakteriocytů, buněk obsahujících symbiotické chemoautotrofní bakterie. Trofosom trávicí trakt funkčně nahrazuje.[51]
V rámci neodermátů aktivně přijímají potravu všechny skupiny vyjma zmiňovaných tasemnic. Jednorodí jsou ektoparazité vodních živočichů, živí se buď pokožkou (Monopisthocotylea), anebo krví (Polyopisthocotylea) svých hostitelů.[52] Dospělé motolice jsou parazity vnitřních orgánů, jako jsou například játra, a živí se tkáněmi a tělními tekutinami hostitele; v některých případech vylučují enzymy, jež tkáň hostitele ještě před pozřením natráví.[53] Motolice mají nejčastěji dvě střevní větve, někdy s postranními výběžky.[38]
Nervová soustava a smysly

Nervová soustava ploštěnců již vykazuje proces cefalizace, tj. koncentraci nervových uzlin do hlavové části těla jakožto výsledek bilaterální symetrie a jednosměrného pohybu vpřed. V hlavové části dochází ke vzniku párového ganglia nebo skupiny ganglií. U primitivnějších forem ploštěnek má nervová soustava charakter síťovitého nervového plexu, zatímco u pokročilejších forem dochází ke vzniku tzv. provazcovité nervové soustavy bez tělních ganglií neboli ortogonální; v takovém případě vybíhá z mozkových zauzlin do těla obyčejně šest nervových provazců, jež jsou spojeny příčnými spojkami (komisurami), přičemž na těchto nervových provazcích se již další ganglia neobjevují. Nervová soustava motolic je ortogonální a svou stavbou podobná nervové soustavě některých ploštěnek, z mozkového centra vybíhají mj. až tři páry podélných nervových provazců. V případě tasemnic je hlavní centrum nervové soustavy ve skolexu, odkud vychází nervy inervující rostellum (část hlavičky s příchytnými háčky) a pár hlavních postranních podélných nervů, které procházejí skrze strobilu. V každé proglotidě tyto postranní nervy spojuje příčná komisura. Často se objevují i další podélné nervy.[54][55]
Většina ploštěnek se orientuje pomocí hmatu. Hmatové receptory mají podobu malých štětinek vystupujících z pokožky, jež bývají soustředěny především v přední polovině těla a v okolí hltanu.[55] Nervové podněty mnohým bentickým ploštěnkám umožňují udržovat svou spodní stranu proti podkladu (pozitivní tigmotaxe pro spodní polovinu těla). Ploštěnky, které volně plavou, mohou mít oproti tomu i statocystu, orgán informující tělo o poloze (tj. na bázi geotaxe). Některé druhy se orientují i pomocí reotaxe, což je reakce organismu indukovaná vodními proudy. Významný vliv má orientace na chemické bázi, a to jak pro vystopování potravy, tak pro nalezení partnera. Chemoreceptory leží po stranách hlavové části, někdy, jako u určitých sladkovodních druhů, dochází i k rozvoji výrazných smyslových laloků. Ploštěnky mohou při orientaci na chemické bázi využívat metodu pokus-omyl, tj. začnou se pohybovat jedním směrem a postupně se snaží měnit směr pomocí slábnoucích/sílících chemických signálů. Jiné ploštěnky, jako rod Dugesia, pohybují přídí ze strany na stranu, aby tak snáze určily směr, odkud nejsilnější chemické signály přicházejí.[56] U ploštěnek se objevují i jednoduché zrakové orgány, takzvaná inverzní očka, pomocí nichž lze však rozeznávat pouze intenzitu světla.[25] Většina ploštěnek vykazuje tzv. negativní fototaxi, a zdržuje se tedy primárně ve tmě.[56] Pár rudimentárních oček mívají i žábrohlísti. Značné redukce doznaly smyslové orgány dospělých endoparazitických druhů, přičemž jsou vesměs omezeny na hmatové receptory (přísavky motolic, skolex tasemnic) a případně i chemoreceptory.[55] Některá jejich vývojová stádia však mohou mít smyslové orgány rozvinutější; například miracidium, invazní stádium motolic, má na rozdíl od dospělé motolice oči[57] a významnou roli při hledání mezihostitelů pro něj mají chemoreceptory.
Rozmnožovací soustava

Rozmnožovací soustava ploštěnců je složena z gonád homologizovatelných s reprodukčními orgány dalších živočichů. Většina ploštěnců je hermafroditická, avšak jedinci mívají oddělené pohlavní orgány.[43] Samčími gonádami jsou varlata (jedno, dvě, anebo i více jako u mnohovětevných ploštěnek), jejichž obsah je odváděn spermatickými kanálky, přičemž se často může hromadit v zásobních semenných váčcích. Spermie mnohdy doplňují sekrety vylučované z přídatných prostatických žlázek. Vlastním kopulačním orgánem, pomocí něhož je vylučován výsledný ejakulát, může být buď penis, anebo tzv. cirrus.[58] Spermie jsou dvoubičíkaté,[59] u některých skupin (řetěznatky, rovnostřevní, neodermáti) existují vysoce modifikované spermie bez bičíků.[60]
Samičí pohlavní soustava je složitější a vyjma vaječníků obsahuje i množství přídatných žláz a struktur, které umožňují například po kopulaci déle uchovávat spermie, anebo naopak přebytečné spermie odvádět. Velká část variací samičích pohlavních orgánů souvisí s tím, jaká ploštěnec vytváří vajíčka. Takzvaná vajíčka endolecitální obsahují větší množství žloutku a nepotřebují další výživu od rodiče, zatímco vajíčka ektolecitálního typu vyžadují druhotnou přítomnost výživných neboli žloutkových žláz. Na základě této charakteristiky lze ploštěnce řadit do dvou velkých skupin: Archoophora a Neoophora (mj. zástupci kladu Neodermata).[43] Rozmnožovací orgány ústí do pohlavního atria, přičemž vývod pro samčí a samičí reprodukční orgány může být buď jednotný, anebo oddělený.[54] Rýhování vajíček u ploštěnců probíhá totálně, spirálním způsobem.[43] Klad Acentrosomata, zahrnující mj. trojvětevné ploštěnky a neodermátní zástupce, ztratil jak spirální rýhování, tak dokonce i centrozomy.[12]
Rozmnožování ploštěnek

Reprodukční biologie ploštěnců je obecně velmi komplikovaná a závisí mj. na tom, jestli jde o volně žijící zástupce, tj. ploštěnky, anebo zástupce parazitických tříd, u nichž se mohou objevovat i složité reprodukční cykly (viz níže). Pro volně žijící ploštěnky je typické vzájemné (křížové) oplodnění, kdy dochází k reciproční výměně spermatu. Další cestou je hypodermické (traumatické) oplodnění, kdy jedna ploštěnka, zastávající úlohu „samce“, probodne svým specializovaným kopulačním orgánem tělní stěnu „samice“ a ejakuluje přímo do mezenchymu.[61] U některých ploštěnek jsou spermie dokonce schopny pronikat přímo skrze pokožku, jako u Thysanozoon nigropapillosum.[62]
U jednostranně se oplodňujících druhů může docházet ke vzniku sexuálních konfliktů, protože je z pohledu evoluční biologie výhodnější být dárcem spermatu než jeho příjemcem. U některých ploštěnek, jako u zástupců rodu Pseudobiceros, vedl tento tlak k rozvoji propracovaného rozmnožovacího chování: pokud se dvě ploštěnky potenciálně připravené k rozmnožování setkají, začnou „šermovat“ svými penisy a snaží se oplodnit druhého jedince. Samooplození se u ploštěnek může objevovat také, nicméně jde o méně častou možnost vzhledem k hrozícímu rychlému úbytku heterozygotnosti během generací. Vyjma hermafroditů existují i gonochorističtí zástupci ploštěnců, jako volně žijící ploštěnky Kronborgia spp. (u kronborgií se navíc objevuje výrazná pohlavní dvojtvárnost z hlediska velikosti).[61]
Snůšku ploštěnky většinou kladou do kokonů.[63] U ploštěnek může být vývoj buď přímý, anebo probíhá přes pelagickou larvu. U velkého množství mnohovětevných ploštěnek se objevuje tzv. Müllerova larva, jež plave pomocí osmi obrvených chlopní a teprve později se usadí na dně, kde se z ní vyvine dospělec. Pro parazitické ploštěnky rodu Stylochus je typický jiný druh larvy, takzvaná Götteho larva, která se vyznačuje pouze čtyřmi takovými chlopněmi.[64]
Vyjma pohlavního způsobu rozmnožování se ploštěnci mohou rozmnožovat i nepohlavně, a to buďto partenogeneticky, anebo pomocí fragmentace samotného těla.[61] Zvláště ploštěnky jsou známé svou vysokou schopností regenerace, již zajišťují nediferencované kmenové buňky zvané neoblasty. Ty tvoří asi pětinu a více buněk dospělého živočicha a jsou zodpovědné za vznik všech dalších typů buněk, přitom tyto diferencované tělní buňky již nevykazují mitotickou aktivitu.[65] Nepohlavní rozmnožování příčným dělením je typické pro sladkovodní a suchozemské druhy ploštěnek. Řetěznatky provádějí mnohonásobné příčné dělení, přičemž jednotliví jedinci zůstávají navzájem spojení v řetězci, dokud nedorostou natolik, aby mohli přežít samostatně.[66] Volně žijící ploštěnky mohou přecházet mezi pohlavním a nepohlavním rozmnožováním, a to v závislosti na okolním prostředí; v méně příznivých podmínkách je z evolučního pohledu obecně výhodnější produkovat geneticky variabilnější potomstvo skrze pohlavní rozmnožování.[61]
V laboratorním prostředí při rozetnutí některých ploštěnek na kusy dochází k regeneraci v nové jedince, různých manipulačních pokusů bylo z tohoto pohledu provedeno velké množství. V místě rány dochází k proliferaci a vytvoření blastému, jenž se diferencuje na nové tkáně. Fragmenty jsou obecně schopny díky Wnt signalizaci udržovat předozadní (anteriorně-posteriorní) osu těla, a regenerovat tak hlavovou část v přední oblasti.[65] Záleží však i na velikosti a vzdálenosti fragmentu od středu těla. Například při rozetnutí ploštěnky v polovině dochází k regeneraci obou polovin, zatímco při odtětí malého kousku zadní části již tento fragment nemusí regenerovat.[67] Řez podél osy přídě může způsobit, že ploštěnce dorostou dvě hlavové části.[65]
Životní cykly kladu Neodermata

Parazitičtí zástupci z kladu Neodermata mají ve srovnání s volně žijícími ploštěnkami mnohem složitější životní cykly.
Nejjednodušší životní a rozmnožovací cyklus lze pozorovat u jednorodých. Jejich jméno vychází z toho, že během svého života využívají pouze jednoho hostitele, přičemž nejčastěji jsou vnějšími parazity na žábrách mořských i sladkovodních ryb, což vysvětluje jejich alternativní jméno žábrohlísti. Některé druhy nicméně mohou parazitovat v průběhu života na vícero orgánech hostitelských druhů, dokonce i endoparaziticky. Za zmínku nicméně stojí rozmnožování žábrohlísta rodu Diplozoon; tento rybí ektoparazit se vyznačuje tím, že pár jedinců navzájem přirůstá pohlavními otvory a takto srostlý zůstává do konce života. Jde o jeden z extrémních případů monogamie v živočišné říši.[61][39]
Endoparazitické třídy motolic a tasemnic mají naproti žábrohlístům velmi složité životní cykly, které zahrnují několik různých mezihostitelů a často dochází ke kombinování nepohlavního a pohlavního rozmnožování v různých fázích životního cyklu. Obecně platí, že takto parazité využívají výhody rychlého množení nepohlavním způsobem v mezihostiteli se zachováním genetické rozmanitosti dané pohlavním rozmnožováním.[61] Obrovského rozvoje dosáhla samotná rozmnožovací soustava. U dospělých motolic zahrnují pohlavní orgány většinu těla (motolice mají oddělená varlata a vaječníky a mohutně rozvinuté žloutkové žlázy) a výsledná vajíčka jsou velmi odolná. Rozmnožování dospělců probíhá nejčastěji pohlavním křížovým oplodněním mezi vícero jedinci obývajícími orgány hostitele, není to však povinností.[40][68] Tasemnice mají v každém článku hrozny drobných varlat, samičí rozmnožovací soustavu tvoří vaječníky a mohutně rozvinuté přídatné žlázy, včetně žláz žloutkových. Vyvíjí se také tzv. děloha (uterus), jež sdružuje vajíčka a postupně může vyplňovat prakticky celou proglotidu. Vajíčka dospělá tasemnice nejčastěji odlučuje přímo s článkem,[41] u malého množství druhů jsou vylučována jednotlivě.[69] Tasemnice se mohou rozmnožovat skrze vzájemné křížové oplodnění, pro zástupce početné skupiny „pravých tasemnic“ Eucestoda (tasemnice šestiostné) je typické samooplození, a to buď v rámci jedné proglotidy, anebo i mezi jednotlivými články jednoho jedince.[70] Rozmnožovací potenciál obou skupin je enormní; některé motolice produkují až 24 000 vajíček za den a některé tasemnice i 720 000. Vzhledem k obrovskému množství potomků, z nichž se však jen část dožije pohlavní dospělosti, může přírodní výběr probíhat velmi efektivně, s velmi rychlou evolucí adaptivních znaků.[71]

Dvojrodé motolice mívají ve svém životním cyklu dva až tři hostitele, přičemž jedním z nich bývá vždy měkkýš, výjimečně mnohoštětinatec. Z obecného hlediska dospělé motolice v definitivním hostiteli produkují vajíčka, která opouštějí jeho tělo například s výkaly, anebo s močí. Vajíčka může následně pozřít mezihostitel, či se z nich líhne vodní pohyblivá larva miracidium, jež mezihostitele – specifického plže – aktivně vyhledá a pronikne do jeho těla. Zde probíhá nepohlavní rozmnožování larev, zahrnující několik různých tělních plánů (měchýřkovitá sporocysta, z jejíchž buněk vznikají redie a následně cerkarie; ačkoli stádium redie může u některých motolic chybět). Plž může být pozřen konečným hostitelem (např. u čeledi Leucochloridiidae), v drtivé většině případů jej však opouští cerkarie v podobě pohyblivých vodních larev, jejichž další vývoj může být různý. U velkého množství motolic vyhledávají dalšího mezihostitele, do něhož pronikají buď aktivně, anebo jsou pozřeny, např. ve slizové kouli. Druhým mezihostitelem již nemusí být výhradně plž, ale – v závislosti na druhu motolice – široké spektrum živočichů. V tkáních druhého mezihostitele se zapouzdřují neboli encystují metacerkarie, které čekají na pozření definitivním hostitelem, ale u některých motolic, např. z čeledí Diplostomatidae, zůstávají cerkarie volné a vytvářejí přechodné stádium mezocerkarie, které se mohou navíc kumulovat v paratenických hostitelích (tj. fakticky jde o čtyřhostitelský cyklus). V případě motolic s dvouhostitelskými cykly jsou některé cerkarie schopny pronikat do definitivního hostitele aktivní cestou, jako v případě krevniček, anebo se zapouzdřují na vegetaci jako metacerkarie a jsou konečným hostitelem pozřeny tímto způsobem (např. čeleď Fasciolidae). Zapouzdřené cerkarie na vegetaci se někdy označují jako adoleskarie.[72][73][74]
Typickým, hojně citovaným příkladem je cyklus motolice jaterní (Fasciola hepatica). Jejich definitivními hostiteli jsou ovce nebo jiní kopytníci. Mezihostitelem je rod sladkovodních plžů bahnatek rodu Lymnaea. Konečné pohyblivé invazivní cerkarie se vyvíjejí přes sporocystu a redii, opouštějí plže skrze jeho plášťovou dutinu a snaží se encystovat na příbřežní vegetaci. Po spasení trávy se cysty dostanou do trávicího traktu definitivního hostitele, kde probíhá jejich vývoj v dospělou motolici.[57] Za zmínku stojí i životní cyklus krevniček (Schistosoma), z nichž některé, jako krevnička močová (S. haematobium), mohou napadat i člověka. Krevničky jsou gonochoristé, přičemž delší a útlejší samice je zachycena v břišní rýze samce. Mezihostiteli jsou různé druhy plžů, pro druh S. haematobium to jsou bulíny (Bulinus). Cerkarie následně aktivně pronikají do pokožky a pak i tělního oběhu definitivního hostitele. Dospělé motolice se lokalizují v různých částech těla, pro krevničku močovou jsou to cévy močového měchýře. Vajíčka jsou hrotitá, rozrušují stěny cév a s krvavou močí jsou vylučována ven z těla.[75][76]

Dospělé tasemnice většinou parazitují v trávicím traktu obratlovců, přičemž obyčejně vyžadují jednoho až dva mezihostitele. Některé tasemnice však mohou využívat i paratenických hostitelů a několik málo druhů má jen jednoho hostitele (např. zástupci rodů Archigetes a Hymenolepis).[77] Životní cykly tasemnic mohou být dosti variabilní, v některých případech je vývoj tasemnic vázán na vodní prostředí, v jiných nikoli. Tasemnice bezbranná (Taenia saginata) a tasemnice dlouhočlenná (T. solium), jejichž definitivním hostitelem je člověk, demonstrují rozmnožovací cyklus tasemnic ze řádu kruhovek, který probíhá výlučně suchozemsky. Mezihostitelem je skot, resp. prase. Oplozená vajíčka, resp. proglotidy odcházejí s výkaly definitivního hostitele do volného prostředí. Ze zygoty se vyvíjí larva onkosféra se šesti larválními háčky, která je chráněna odolným embryoforem, tj. relativně pevnou, nepropustnou obalovou vrstvou zárodku tvořenou proteiny podobnými keratinu. Po pozření vajíček mezihostitelem proniká onkosféra do různých částí těla (svalstvo, vnitřní orgány) a mění se v sekundární mechýřkovitou larvocystu neboli boubel s vchlípeným skolexem. Konzumací masa s larvocystami se nakazí definitivní hostitel. Pro tasemnici bezbrannou a dlouhočlennou je typická larvocysta typu cysticercus, s jedním skolexem v měchýřku, pro jiné druhy kruhovek jsou typické jiné typy larvocyst (cysticercoid, coenurus nebo echinokok).[78][79] Tasemnice ze řádu štěrbinovek naproti tomu k vývoji vyžadují vodní prostředí, příkladem je škulovec široký (Dibothriocephalus latus). Ve vodě se z vajíček škulovce líhnou obrvené larvy koracidia. Koracidium musí být pozřeno klanonožcem, který představuje prvního mezihostitele. Sekundárním mezihostitelem je ryba, definitivním hostitelem rybožraví savci včetně člověka. Vývojová stádia v klanonožci a rybě se nazývají procerkoid a plerocerkoid.[78]
 Cyklus motolice jaterní
Cyklus motolice jaterní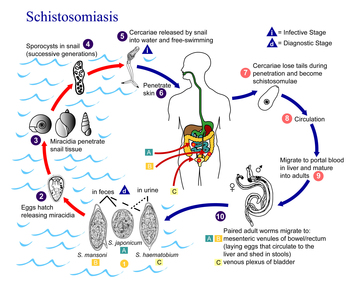 Cyklus schistosom
Cyklus schistosom Cyklus tasemnice bezbranné
Cyklus tasemnice bezbranné Cyklus škulovce širokého
Cyklus škulovce širokého
Rozšířený fenotyp a parazitární manipulace

Vzhledem k tomu, že ploštěnci z kladu Neodermata využívají ke svým rozmnožovacím cyklům větší množství hostitelských druhů, často vyvinuli rozličné postupy, jak hostitele co nejvíce využít ve svůj prospěch, ať už z hlediska větší prosperity samotného parazita během jeho vývoje, nebo ke snazšímu proniknutí do trávicího traktu definitivního hostitele. Jde o příklad rozšířeného fenotypu, který lze demonstrovat na velkém množství parazitických druhů.
Například larvy motolic nutí po infikování plže, aby budovali silnější schránky, a poskytovali jim tak bezpečné útočiště pro jejich vlastní rozmnožování. Motolice jsou také schopny cílené parazitické kastrace mezihostitelských plžů.[80] Tím, že parazit znemožní reprodukci hostitelského organismu, přesměruje jeho energetické zdroje na růst a regeneraci, čímž zajistí jeho delší přežívání. Například infekce krevničkou střevní (Schistosoma mansoni) vede u mezihostitelských okružáků (Biomphalaria) k poklesu hladiny serotoninu v krvi i plazmě, přičemž hladina tohoto monoaminu výrazně koreluje s produkováním vajíček okružáka. Hormonální změny jiného účelu lze pozorovat u tasemnice Taenia crassiceps, jejíž larvy infikují myšovité hlodavce. Infekce samců vede k výraznému poklesu samčích pohlavních hormonů a naopak až k 200násobné produkci estradiolu; samice jsou totiž vůči infekci ve srovnání se samci mnohem méně rezistentní a feminizace samců ze strany parazita tuto samčí výhodu stírá.[71][81]
Jiné parazitární manipulace přímo napomáhají ulovení jedince, aby tak parazit mohl snáze přejít do dalšího hostitele. Například tasemnice rodu Schistocephalus či motolice Diplostomum spathaceum mají jako mezihostitele ryby, přičemž v obou případech jimi manipulují tak, aby plavaly blíže k hladině a staly se tak kořistí rybožravých ptáků: ryby nakažené tasemnicí vyžadují více kyslíku, zatímco motolice parazitují v očních čočkách ryb a zakalují jim zrak.[80] Propracovaný případ parazitární manipulace lze pozorovat u plžů jantarek infikovaných motolicí podivnou (Leucochloridium paradoxum). Sporocysty motolice infikují tykadla plže, která se zvětší, pestře zbarví a začnou pulzovat, svým vzhledem tak připomínají housenku. Plž se zároveň začíná vyhýbat skrytým místům, a stává se tak snadnou kořistí ptáků – definitivních hostitelů.[74] Cerkarie motolice kopinaté (Dicrocoelium dendriticum) jsou zase parazity mravenců. Definitivním hostitelem je ovce, jež musí mravence pozřít, motolice tedy nutí svého sekundárního mezihostitele, aby zůstal – vyjma nejteplejších částí dne – zakousnutý do listu trávy a čekal zde na svůj osud.[72][80] Motolice kopinatá zároveň demonstruje příklad tzv. pseudoaltruismu. Její cerkarie, vznikající nepohlavním rozmnožováním, opouštějí tělo plže a následně jsou mravencem pozřeny společně (v jakési slizové kouli), přičemž jedna cerkarie se usadí v ganglionu mravence a ostatní se opouzdří v jeho těle. Larva v ganglionu jako jedinec uhyne, ale manipulací s hostitelem pomáhá prosadit se ostatním larvám – svým genetickým kopiím.[71]
Diverzita a výskyt

Je známo asi 26 500 druhů ploštěnců, z toho asi 6 500 druhů volně žijících ploštěnek, 8 000 druhů tasemnic a 12 000 druhů motolic.[82] Velké množství druhů však stále pravděpodobně zůstává nepopsáno. Počet nepopsaných ploštěnek je odhadován na možná několik desítek tisíc, odhadovaný počet všech neodermátů pak na 40 000 až 100 000 druhů.[10]
Ploštěnky se vyskytují prakticky po celém světě, většinou v mořském a sladkovodním prostředí. Mohou být extrémně hojné v rámci mořské meiofauny a obyčejně představují důležité predátory ekosystému. Mnoho drobných druhů žije vmezeřeno mezi zrnky písku ve vodních stanovištích, větší ploštěnky se pohybují na dně, anebo mohou plavat pelagicky. Některé ploštěnky byly nalezeny i v oblasti Peruánsko-chilského či Kurilského příkopu. Trojvětevné ploštěnky mohou žít suchozemsky, často ve vlhkém listí a půdě. Zmínit lze například rod Obama, jenž je původní v jihoamerickém atlantickém lese.[56][83] Při zavlečení do nepůvodních areálů se mohou ploštěnky stávat invazními druhy. Jedním z nich je například ploštěnka novozélandská (Arthurdendyus triangulata, syn. Artioposthia triangulata), jež původně žila na Novém Zélandu a představuje evropský invazní druh, který se svou predací výrazně podepisuje na populaci žížal.[84][85]
Ještě rozšířenější jsou zástupci kladu Neodermata, jejichž evoluční úspěch je výsledkem kombinace vzniku tegumentu a schopností parazitovat na obrovském množství druhů hostitelských živočichů. Většina druhů obratlovců, nehledě k mnoha mezihostitelům z různých živočišných kmenů, představuje hostitele pro alespoň jeden druh parazitického ploštěnce.[10] Především tasemnice byly nalezeny v širokém spektru obratlovců i napříč různými extrémními životními prostředími. Asi třetina tasemnic představuje parazity ryb a paryb, z toho některé štěrbinovky byly objeveny i u hlubokomořských rybích druhů ze severního Atlantiku žijících v hloubce téměř 2 300 metrů. Více než stovka druhů tasemnic byla popsána z ptačích a savčích druhů z arktických a antarktických biotopů.[69]
Parazitičtí ploštěnci a člověk

Tasemnice a motolice vyvolávají nemoci u lidí, resp. jejich hospodářských zvířat. Významné ekonomické ztráty v pastevních chovech ovcí a dobytka může působit například motolice jaterní (Fasciola hepatica) z čeledi Fasciolidae; do stejné čeledi pak spadá i původně severoamerická motolice Fascioloides magna, proslavená svou vysokou patogenitou pro evropské přežvýkavce, kteří na ni nejsou adaptováni.[86] Z patogenních tasemnic lze naopak jmenovat zástupce čeledi Anoplocephalidae, již platí za významný faktor v etiologii kolikových onemocnění koní.[87] Opomenut nemůže být ani vliv žábrohlístů, kteří mohou způsobit vážné ztráty populací na rybích farmách (např. Gyrodactylus salaris).[88]
Člověk je typickým definitivním hostitelem některých motolic rodu Schistosoma (S. mansoni, S. haematobium a S. japonicum) a tasemnic Taenia saginata, T. solium a T. asiatica. U ostatních ploštěnců představuje atypického, resp. jednoho z možných definitivních hostitelů nebo mezihostitelů.
Schistosomóza (vyvolávaná krevničkami) nebo také bilharzióza je po malárii druhé nejčastější parazitární onemocnění člověka. Podle WHO postihuje téměř 240 milionů lidí na celém světě a více než 700 milionů lidí žije v endemických oblastech,[89] především v Africe, Asii, Jižní Americe a na některých karibských ostrovech. Průnik cerkarií do krevního oběhu člověka může vést k akutním nebo chronickým infekcím, přičemž chronický průběh onemocnění je u krevničky močové spojen s rizikem rozvoje rakoviny močového měchýře.[75] Na člověku může parazitovat mj. zmíněná motolice jaterní, infekce encystovanými cerkariemi je spojena s napadením žlučovodů a následnými nekrotickými a zánětlivými reakcemi. Především v pevninské jihovýchodní Asii hrozí infekce motolicí žlučovou (Clonorchis sinensis) a motolicí thajskou (Opisthorchis viverrini), pro něž představují sekundární mezihostitele kaprovité ryby, nebezpečí tedy hrozí ze strany nedostatečně tepelně opracovaného rybího masa.[90] K roku 2012 bylo jenom motolicí žlučovou infikováno asi 15 až 20 milionů lidí.[91]
K roku 2000 bylo odhadem nakaženo 45 milionů lidí tasemnicí bezbrannou a 3 miliony lidí tasemnicí dlouhočlennou.[90] V případě tasemnice dlouhočlenné člověk figuruje jako definitivní hostitel, kdy infekce nevykazuje vážnější průběh, nicméně může se nakazit i jejími vajíčky, což následně vede k rozvoji cysticerkózy, závažného onemocnění vyvolané rozvojem larvocyst tasemnice. Neurocysticerkóza, při níž larvocysty napadnou mozek, je celosvětově nejčastější příčinou získané epilepsie a v lokalitách, kde se parazit vyskytuje endemicky, způsobuje 30 % všech případů epilepsie.[92] Zajímavé je, že tasemnice rodu Taenia se zřejmě staly parazity lidí ještě před rozvojem zemědělství a domestikací hospodářských zvířat, a to již někdy na přelomu pliocénu a pleistocénu, kdy se tehdy žijící příslušníci rodu Homo začali živit masem a následně se na ně adaptovaly druhy tasemnic původně přizpůsobené infekci kočkovitých či hyenovitých šelem.[93]
Člověk je atypickým mezihostitelem také pro tasemnice rodu Echinococcus, jež vyvolávají echinokokózu. Například měchožil zhoubný (E. granulosus) v takovém případě vytváří mohutné larvocysty, z nichž se po prasknutí uvolňují zárodky vzniklé nepohlavním rozmnožováním, které se roznášejí po těle a dávají vznik dalším larvocystám. U člověka může tato situace vést k fatální alergické šokové reakci. Velmi závažnou formu echinokokózy vyvolává infekce měchožilem bublinatým (E. multilocularis), jehož larvální stádium nevytváří solitérní cystu, ale prorůstá do okolních tkání a imituje nádorové bujení.[32][94]
Odkazy
Poznámky
- ↑ Břichobrvky byly tradičně sdružovány k hlístům, moderní studie je sdružují s ploštěnci.[14] Zároveň je pravděpodobné, že blízce příbuzné ploštěncům a břichobrvkám jsou rovněž sépiovky, původně klasifikované jako součást nepřirozené skupiny morulovců. Pokud břichobrvky a sépiovky tvoří sesterské taxony, ploštěnci vystupují jakožto sesterský taxon vůči celému tomuto kladu.[15]
- ↑ Vrtejši jsou považováni za vnitřní skupinu vířníků, v takovém případě je pro skupinu užíváno jména Syndermata
Reference
- ↑ Brusca, Moore a Shuster 2016, s. 375–376.
- ↑ Hutchins, Thoney a Schlage 2003, s. 11.
- ↑ History of the Classification of Animals [online]. biologydiscussion.com, 2016-05-27 [cit. 2022-03-14]. Dostupné online. (anglicky)
- ↑ Giribet a Edgecombe 2020, s. 92–93.
- ↑ a b Giribet a Edgecombe 2020, s. 88.
- ↑ WESTBLAD, E. Xenoturbella bocki n.g., n.sp., a peculiar, primitive turbellarian type. Arkiv för Zoologi. 1949.
- ↑ Brusca, Moore a Shuster 2016, s. 25.
- ↑ GAISLER, Jiří; ZIMA, Jan. Zoologie obratlovců. 2. vyd. Praha: Academia, 2007. ISBN 978-80-200-1484-9. S. 31–32.
- ↑ Giribet a Edgecombe 2020, s. 326–327.
- ↑ a b c d e LAUMER, Christopher E; HEJNOL, Andreas; GIRIBET, Gonzalo. Nuclear genomic signals of the ‘microturbellarian’ roots of platyhelminth evolutionary innovation. eLife. 2015-03-12, roč. 4, s. e05503. Dostupné online [cit. 2022-03-16]. ISSN 2050-084X. doi:10.7554/eLife.05503.
- ↑ a b PERKINS, Elizabeth M.; DONNELLAN, Steve C.; BERTOZZI, Terry. Closing the mitochondrial circle on paraphyly of the Monogenea (Platyhelminthes) infers evolution in the diet of parasitic flatworms. International Journal for Parasitology. 2010-09-01, roč. 40, čís. 11, s. 1237–1245. Dostupné online [cit. 2022-03-23]. ISSN 0020-7519. doi:10.1016/j.ijpara.2010.02.017. (anglicky)
- ↑ a b EGGER, Bernhard; LAPRAZ, François; TOMICZEK, Bartłomiej. A Transcriptomic-Phylogenomic Analysis of the Evolutionary Relationships of Flatworms. Current Biology. 2015-05-18, roč. 25, čís. 10, s. 1347–1353. PMID 25866392. Dostupné online [cit. 2022-03-16]. ISSN 0960-9822. doi:10.1016/j.cub.2015.03.034. PMID 25866392. (anglicky)
- ↑ Giribet a Edgecombe 2020, s. 327–329.
- ↑ a b Giribet a Edgecombe 2020, s. 311–312.
- ↑ LU, Tsai-Ming; KANDA, Miyuki; SATOH, Noriyuki; FURUYA, Hidetaka. The phylogenetic position of dicyemid mesozoans offers insights into spiralian evolution. Zoological Letters [online]. BioMed Central Ltd, part of Springer Nature, 2017-05-29 [cit. 2021-04-19]. Roč. 3:6. Dostupné online. ISSN 2056-306X. doi:10.1186/s40851-017-0068-5. (anglicky)
- ↑ STRUCK, Torsten H.; WEY-FABRIZIUS, Alexandra R.; GOLOMBEK, Anja. Platyzoan Paraphyly Based on Phylogenomic Data Supports a Noncoelomate Ancestry of Spiralia. Molecular Biology and Evolution. 2014-07-01, roč. 31, čís. 7, s. 1833–1849. Dostupné online [cit. 2022-03-17]. ISSN 0737-4038. doi:10.1093/molbev/msu143.
- ↑ MARLÉTAZ, Ferdinand; PEIJNENBURG, Katja T. C. A.; GOTO, Taichiro. A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans. Current Biology. 2019-01-21, roč. 29, čís. 2, s. 312–318.e3. Dostupné online [cit. 2022-03-17]. ISSN 0960-9822. doi:10.1016/j.cub.2018.11.042. (anglicky)
- ↑ a b c Giribet a Edgecombe 2020, s. 345.
- ↑ a b c Brusca, Moore a Shuster 2016, s. 407.
- ↑ KNAUST, Dirk; DESROCHERS, André. Exceptionally preserved soft-bodied assemblage in Ordovician carbonates of Anticosti Island, eastern Canada. Gondwana Research. 2019-07-01, roč. 71, s. 117–128. Dostupné online [cit. 2022-03-23]. ISSN 1342-937X. doi:10.1016/j.gr.2019.01.016. (anglicky)
- ↑ a b De Baets, K., P. Dentzien-Dias, I. Upeniece, O. Verneau and P. C. J. Donoghue. "Chapter Three – Constraining the Deep Origin of Parasitic Flatworms and Host-Interactions with Fossil Evidence". In Kenneth De Baets and D. Timothy J Littlewood (ed.). Advances in Parasitology. Advances in Parasitology. Vol. 90. pp. 93–135. doi:10.1016/bs.apar.2015.06.002. ISBN 978-0-12-804001-0
- ↑ DENTZIEN-DIAS, Paula C.; POINAR, George; DE FIGUEIREDO, Ana Emilia Q. Tapeworm Eggs in a 270 Million-Year-Old Shark Coprolite. PLoS ONE. 2013-01-30, roč. 8, čís. 1, s. e55007. Dostupné online [cit. 2022-03-18]. ISSN 1932-6203. doi:10.1371/journal.pone.0055007. PMID 23383033. (anglicky)
- ↑ ZRZAVÝ, Jan. Fylogeneze živočišné říše. 1. vyd. Praha: Scientia 255 s. ISBN 80-86960-08-0, ISBN 978-80-86960-08-1. OCLC 124086610
- ↑ a b c d Sedlák 2002, s. 41.
- ↑ a b c d e f Smrž 2015, s. 36.
- ↑ a b c Brusca, Moore a Shuster 2016, s. 385.
- ↑ a b c d Brusca, Moore a Shuster 2016, s. 391.
- ↑ a b Giribet a Edgecombe 2020, s. 329–331.
- ↑ Smrž 2015, s. 37.
- ↑ Phylum Platyhelminthes. www.shapeoflife.org [online]. 2014-02-13 [cit. 2022-03-24]. Dostupné online. (anglicky)
- ↑ Hutchins, Thoney a Schlage 2003, s. 228.
- ↑ a b Sedlák 2002, s. 48.
- ↑ a b c Giribet a Edgecombe 2020, s. 331–332.
- ↑ Nielsen 2012, s. 166.
- ↑ a b Brusca, Moore a Shuster 2016, s. 382–384.
- ↑ LENGERER, Birgit; PJETA, Robert; WUNDERER, Julia. Biological adhesion of the flatworm Macrostomum lignano relies on a duo-gland system and is mediated by a cell type-specific intermediate filament protein. Frontiers in Zoology. 2014-02-12, roč. 11, čís. 1, s. 12. Dostupné online [cit. 2022-03-25]. ISSN 1742-9994. doi:10.1186/1742-9994-11-12. PMID 24520881.
- ↑ Sedlák 2002, s. 43.
- ↑ a b Sedlák 2002, s. 45.
- ↑ a b Smrž 2015, s. 43.
- ↑ a b Smrž 2015, s. 41.
- ↑ a b Smrž 2015, s. 44.
- ↑ a b Giribet a Edgecombe 2020, s. 332–333.
- ↑ a b c d Smrž 2015, s. 37–38.
- ↑ Smrž 2015, s. 39.
- ↑ a b Hutchins, Thoney a Schlage 2003, s. 187–188.
- ↑ RITSON-WILLIAMS, Raphael; YOTSU-YAMASHITA, Mari; PAUL, Valerie J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proceedings of the National Academy of Sciences. 2006-02-21, roč. 103, čís. 9, s. 3176–3179. Dostupné online [cit. 2022-03-27]. ISSN 0027-8424. doi:10.1073/pnas.0506093103. PMID 16492790.
- ↑ Brusca, Moore a Shuster 2016, s. 389.
- ↑ Smrž 2015, s. 40.
- ↑ VENN, A. A.; LORAM, J. E.; DOUGLAS, A. E. Photosynthetic symbioses in animals. Journal of Experimental Botany. 2008, s. 1069–1080. Dostupné online [cit. 2022-03-27].
- ↑ VAN STEENKISTE, Niels W. L.; STEPHENSON, India; HERRANZ, María; HUSNIK, Filip; KEELING, Patrick J.; LEANDER, Brian S. A new case of kleptoplasty in animals: Marine flatworms steal functional plastids from diatoms. Science Advances [online]. American Association for the Advancement of Science, 2019-07-05 [cit. 2022-08-18]. Roč. 5, čís. 7: eaaw4337. Dostupné online. Dostupné také na: [1]. ISSN 2375-2548. doi:10.1126/sciadv.aaw4337. PMID 31328166. (anglicky)
- ↑ LEISCH, Nikolaus; DIRKS, Ulrich; GRUBER-VODICKA, Harald R. Microanatomy of the trophosome region of Paracatenula cf. polyhymnia (Catenulida, Platyhelminthes) and its intracellular symbionts. Zoomorphology. 2011, roč. 130, čís. 4, s. 261–271. PMID 22131640 PMCID: PMC3213344. Dostupné online [cit. 2022-04-04]. ISSN 0720-213X. doi:10.1007/s00435-011-0135-y. PMID 22131640.
- ↑ Hutchins, Thoney a Schlage 2003, s. 214.
- ↑ Hutchins, Thoney a Schlage 2003, s. 200.
- ↑ a b Sedlák 2002, s. 42.
- ↑ a b c Brusca, Moore a Shuster 2016, s. 392–394.
- ↑ a b c Hutchins, Thoney a Schlage 2003, s. 187.
- ↑ a b Franc 2005, s. 29.
- ↑ Brusca, Moore a Shuster 2016, s. 395–397.
- ↑ Franc 2005, s. 27.
- ↑ Giribet a Edgecombe 2020, s. 338.
- ↑ a b c d e f RAMM, Steven A. Exploring the sexual diversity of flatworms: Ecology, evolution, and the molecular biology of reproduction. Molecular Reproduction and Development. 2016-07-07, roč. 84, čís. 2, s. 120–131. Dostupné online [cit. 2022-03-31]. ISSN 1040-452X. doi:10.1002/mrd.22669.
- ↑ JIE, Wei-Ban; KUO, Shih-Chieh; MOK, Hin-Kiu. Re-description of Thysanozoon nigropapillosum (Polycladida: Pseudocerotidae) from the South China Sea, with observations on a novel pre-copulatory structure, sexual behaviour and diet. RAFFLES BULLETIN OF ZOOLOGY. 2014, s. 764–770. Dostupné online.
- ↑ Smrž 2015, s. 38.
- ↑ Hutchins, Thoney a Schlage 2003, s. 188.
- ↑ a b c Giribet a Edgecombe 2020, s. 335–336.
- ↑ Brusca, Moore a Shuster 2016, s. 394.
- ↑ Brusca, Moore a Shuster 2016, s. 395.
- ↑ HANNA, Robert. Fasciola hepatica: Histology of the Reproductive Organs and Differential Effects of Triclabendazole on Drug-Sensitive and Drug-Resistant Fluke Isolates and on Flukes from Selected Field Cases. Pathogens. 2015-06-26, roč. 4, čís. 3, s. 431–456. PMID 26131614 PMCID: PMC4584266. Dostupné online [cit. 2022-04-02]. ISSN 2076-0817. doi:10.3390/pathogens4030431. PMID 26131614.
- ↑ a b Hutchins, Thoney a Schlage 2003, s. 229.
- ↑ Brusca, Moore a Shuster 2016, s. 403.
- ↑ a b c FLEGR, Jaroslav. Evoluční biologie. 2., opr. a rozš. vyd. vyd. Praha: Academia, 2009. ISBN 978-80-200-1767-3, ISBN 80-200-1767-4. OCLC 505914273 S. 114–115, 343, 359–361.
- ↑ a b HORÁK, P. Motolice – parazitičtí červi s nejkomplikovanějšími životními cykly. ŽIVA AVČR. 2010, čís. 5, s. 230–232. Dostupné online.
- ↑ Volf a Horák 2007, s. 154–156.
- ↑ a b Giribet a Edgecombe 2020, s. 339–342.
- ↑ a b NELWAN, Martin L. Schistosomiasis: Life Cycle, Diagnosis, and Control. Current Therapeutic Research. 2019-01-01, roč. 91, s. 5–9. Dostupné online [cit. 2022-04-02]. ISSN 0011-393X. doi:10.1016/j.curtheres.2019.06.001. (anglicky)
- ↑ Sedlák 2002, s. 46.
- ↑ Volf a Horák 2007, s. 181.
- ↑ a b Brusca, Moore a Shuster 2016, s. 405.
- ↑ Smrž 2015, s. 45.
- ↑ a b c ZRZAVÝ, Jan; MIHULKA, Stanislav; STORCH, David; BURDA, Hynek; BEGALL, Sabine. Jak se dělá evoluce : labyrintem evoluční biologie. 4. vyd. Praha: Dokořán, Argo 479 s. ISBN 978-80-7363-763-7, ISBN 80-7363-763-4. OCLC 982100649 S. 154–156.
- ↑ LARRALDE, C.; MORALES, J.; TERRAZAS, I. Sex hormone changes induced by the parasite lead to feminization of the male host in murine Taenia crassiceps cysticercosis. The Journal of Steroid Biochemistry and Molecular Biology. 1995-06, roč. 52, čís. 6, s. 575–580. PMID 7779761. Dostupné online [cit. 2022-04-02]. ISSN 0960-0760. doi:10.1016/0960-0760(95)00062-5. PMID 7779761.
- ↑ Brusca, Moore a Shuster 2016, s. 376.
- ↑ Giribet a Edgecombe 2020, s. 324–325.
- ↑ BOAG, B.; EVANS, K. A.; YEATES, G. W. Assessment of the global potential distribution of the predatory land planarian Artioposthia triangulata (Dendy) (Tricladida: Terricola) from ecoclimatic data. New Zealand Journal of Zoology. 1995-01-01, roč. 22, čís. 3, s. 311–318. Dostupné online [cit. 2022-04-03]. ISSN 0301-4223. doi:10.1080/03014223.1995.9518046.
- ↑ HENEBERG, Petr. Invaze ploštěnek do Evropy. Když jen dotyk znamená smrt. ŽIVA AVČR. 2008, čís. 3, s. 122–123. Dostupné online.
- ↑ Volf a Horák 2007, s. 164.
- ↑ Vojtková M., Mezerová J., Koudela B. Výskyt a klinický význam tasemnice Anoplocephala perfoliata. Veterinářství 2006;56:24-28. [2]
- ↑ Volf a Horák 2007, s. 177.
- ↑ Schistosomiasis (Bilharzia). www.who.int [online]. [cit. 2022-10-03]. Dostupné online. (anglicky)
- ↑ a b NORTHROP-CLEWES, C. A; SHAW, C. Parasites. British Medical Bulletin. 2000-01-01, roč. 56, čís. 1, s. 193–208. Dostupné online [cit. 2022-04-03]. ISSN 0007-1420. doi:10.1258/0007142001902897. (anglicky)
- ↑ HONG, Sung-Tae; FANG, Yueyi. Clonorchis sinensis and clonorchiasis, an update. Parasitology International. 2012-03-01, roč. 61, čís. Opisthorchiasis and clonorchiasis: Major neglected tropical diseases in Eurasia, s. 17–24. Dostupné online [cit. 2022-04-13]. ISSN 1383-5769. doi:10.1016/j.parint.2011.06.007. (anglicky)
- ↑ Taeniasis/Cysticercosis. www.who.int [online]. WHO, 11 January 2022 [cit. 2022-04-03]. Dostupné online. (anglicky)
- ↑ HOBERG, E. P.; ALKIRE, N. L.; QUEIROZ, A. D. Out of Africa: origins of the Taenia tapeworms in humans. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2001-04-22, roč. 268, čís. 1469, s. 781–787. Dostupné online [cit. 2022-04-03]. ISSN 0962-8452. doi:10.1098/rspb.2000.1579. PMID 11345321. (anglicky)
- ↑ Volf a Horák 2007, s. 189–190.
Literatura
- Česky
- VOLF, Petr; HORÁK, Petr, 2007. Paraziti a jejich biologie. Praha: Triton. ISBN 9788073870089, ISBN 8073870088.
- SEDLÁK, Edmund, 2002. Zoologie bezobratlých. Brno: Přírodovědecká fakulta MU. ISBN 80-210-2892-0.
- SMRŽ, Jaroslav, 2015. Základy biologie, ekologie a systému bezobratlých živočichů. Praha: Karolinum Press. ISBN 8024622580, ISBN 9788024622583.
- Slovensky
- FRANC, V, 2005. Systém a fylogenéza živočíchov – bezchordáty. 2. vyd. Banská Bystrica: Univerzita Mateje Bela. (slovensky)
- Anglicky
- BRUSCA, R. C.; MOORE, Wendy; SHUSTER, S. M, 2016. Invertebrates. Sunderland, Massachusetts: Sinauer Associates, lnc., Publishers. Dostupné online. ISBN 9781605353753. (anglicky)
- GIRIBET, Gonzalo; EDGECOMBE, Gregory, 2020. The Invertebrate Tree of Life. Princeton, NJ Oxford: Princeton University Press. ISBN 978-0-691-19706-7, ISBN 0-691-19706-7. OCLC 1129197548 Kapitola 43: PLATYHELMINTHES. (anglicky)
- NIELSEN, Claus, 2012. Animal Evolution : Interrelationships of The Living Phyla. 3. vyd. Oxford: OUP. Dostupné online. ISBN 978-0-19-162530-5, ISBN 0-19-162530-2. Kapitola 34: Phylum Platyhelminthes. (anglicky)
- HUTCHINS, Michael; THONEY, Dennis A.; SCHLAGE, Neil, 2003. Lower Metazoans And Lesser Deuterostomes. 2. vyd. Farmington Hills, MI: Gale. (Grzimek's Animal Life Encyclopedia; sv. 1). ISBN 0787657778, ISBN 9780787657772. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu ploštěnci na Wikimedia Commons
Obrázky, zvuky či videa k tématu ploštěnci na Wikimedia Commons  Taxon Platyhelmnithes ve Wikidruzích
Taxon Platyhelmnithes ve Wikidruzích- Seznam děl v Souborném katalogu ČR, jejichž tématem jsou ploštěnci
- BioLib.cz – Platyhelminthes (Ploštěnci) [online]. BioLib.cz. Dostupné online.
- ROČEK, Zbyněk. Obecná morfologie živočichů [online]. Dostupné online.
- MYERS, P. Platyhelminthes [online]. Animal Diversity Web, 2002 [cit. 2022-04-14]. Dostupné online. (anglicky)
- BERGEY, E. A.; BRIGHT, E. G. Phylum Platyhelminthes [online]. University of Oklahoma [cit. 2022-04-14]. Dostupné online. (anglicky)
Média použitá na této stránce

Autor:
- Information-silk.png: Mark James
- derivative work: KSiOM(Talk)
{kind=link}
A tiny blue 'i' information icon converted from the Silk icon set at famfamfam.com
Autor: Gieysztoria, Licence: CC BY-SA 4.0
Ploštěnka Dolichomacrostomum uniporum z infrakmene Macrostomorpha
Autor: Alex Lin and Bret Pearson., Licence: CC BY 3.0
Distribuição de Neoblastos pelo corpo de uma planária da espécie Schmidtea mediterranea. Os Neoblastos aparecem pelo corpo todo com exceção da faringe mostrada por um seta. Pontos verdes são Neoblastos que estão se dividindo enquanto pontos vermelhos são Neoblastos que não estão se dividindo
(c) Betty Wills, CC BY-SA 4.0
Yellow papillae flatworm (Thysanozoon nigropapillosum) swimming at 40 ft (12 m) depth, Manta Ray Bay, Yap, Federated States of Micronesia.
Autor: The Other 95%, Licence: CC BY-SA 4.0
Head, auricles, ocelli, rami, and pharynx

Autor: Andreas Neudecker, Licence: CC BY-SA 4.0
Dugesia gonocephala, schematic, showing pigment-cup ocelli, nervous system, digestive system
Autor: Stefan Walkowski, Licence: CC BY-SA 4.0
Vajíčko tasemnice bezbranné, vnitřní embryofor s viditelnou onkosférou uprostřed
Diplozoon paradoxum (Nordmann) = Diplozoon paradoxum von Nordmann, 1832, mated pair from below

life cycle of en:Diphyllobothrium latum.
It's from the CDC (US government), so it's not copyrighted.Autor: Flukeman, Licence: CC BY-SA 3.0
Adult of Fascioloides magna isolated from liver of red deer
Autor: Jean-Lou Justine, Leigh Winsor, Delphine Gey, Pierre Gros, Jessica Thévenot, Licence: CC BY-SA 4.0
Figure 6 from Article
Obama nungara, dark form feeding on an earthworm. The everted pharynx can be clearly seen partly enveloping the head of the earthworm (unidentified species). Specimen MNHN JL092 from Montauban, Tarn-et-Garonne. Photo by Pierre Gros.
Autor: John Alan Elson, Licence: CC BY-SA 4.0
Planaria Cross Section Magnified 40 times
Autor: Stephen Childs, Licence: CC BY 2.0
Barevná mořská ploštěnka Pseudoceros liparus pochází z tropického Indopacifiku
Magnetic resonance image of a patient with neurocysticercosis demonstrating multiple cysticerci within the brain.
Autor:
| This image is created by user Dick Belgers at Waarneming.nl, a source of nature observations in the Netherlands. |
Jantarka obecná (Succinea putris) nakažená motolicí podivnou (Leucochloridium paradoxum)
Autor: Russ Hobbs, Klaus Rohde, Licence: CC BY-SA 3.0
Adult Gigantolina elongata in the body cavity of a long-necked turtle, Chelodina longicollis
Autor: Circa24, Licence: CC BY-SA 3.0
Attachment organ (Haptor or opisthaptor) of the Monogenea (genus: Cleiodiscus).
Autor: Photo courtesy of Nico Michiels., Licence: CC BY 2.5
Two flatworms of the genus Pseudobiceros bedfordi about to engage in a sperm battle. They are hermaphroditic and have two penises that are used to inject sperm into the partner. P. bedfordi is exceptional in that it applies sperm onto the partner's skin, rather than injecting it.
Autor: Flukeman, Licence: CC BY-SA 4.0
Fasciola hepatica cercariae emerged from infected snail Galba truncatula
A flame cell and tube cell, the excretory system of flatworms

Eggs are eliminated with feces or urine (1). Under optimal conditions the eggs hatch and release miracidia (2), which swim and penetrate specific snail intermediate hosts (3). The stages in the snail include 2 generations of sporocysts (4) and the production of cercariae (5). Upon release from the snail, the infective cercariae swim, penetrate the skin of the human host (6), and shed their forked tail, becoming schistosomulae (7). The schistosomulae migrate through several tissues and stages to their residence in the veins (8,9). Adult worms in humans reside in the mesenteric venules in various locations, which at times seem to be specific for each species (10). For instance, S. japonicum is more frequently found in the superior mesenteric veins draining the small intestine [A], and S. mansoni occurs more often in the superior mesenteric veins draining the large intestine [B]. However, both species can occupy either location, and they are capable of moving between sites, so it is not possible to state unequivocally that one species only occurs in one location. S. haematobium most often occurs in the venous plexus of bladder [C], but it can also be found in the rectal venules. The females (size 7 to 20 mm; males slightly smaller) deposit eggs in the small venules of the portal and perivesical systems. The eggs are moved progressively toward the lumen of the intestine (S. mansoni and S. japonicum) and of the bladder and ureters (S. haematobium), and are eliminated with feces or urine, respectively (1).
ID#: 5260
Description: This is an adult Taenia saginata tapeworm.
Humans become infected by ingesting raw or undercooked infected meat. In the human intestine, the cysticercus develops over 2 mo. into an adult tapeworm, which can survive for years, attaching to, and residing in the small intestine.
Content Providers(s): CDC Creation Date: 1986
Copyright Restrictions: None - This image is in the public domain and thus free of any copyright restrictions. As a matter of courtesy we request that the content provider be credited and notified in any public or private usage of this image.
The life cycle of Taenia saginata