Stonek

Z uzlin vyrůstají řapíky listů a postranní větve
Stonek je vegetativní orgán cévnatých rostlin a společně s kořeny a listy představuje hlavní stavební jednotku rostlinného těla. Charakter stonku a způsob jeho větvení určuje růstovou formu rostliny, tedy jedná-li se o strom, keř, bylinu, liánu a podobně. Stonek nese nad zemí listy a reprodukční orgány (květy a plody), pod zemí kořeny. Mimo to může plnit i řadu různých dalších funkcí. U většiny jednoděložných rostlin je stonek bylinný, u jehličnanů a většiny dvouděložných je druhotně tloustnoucí a více či méně dřevnatý.
Funkce
Stonek je základním orgánem velké většiny cévnatých rostlin. Jako takový má i některé zásadní funkce. Nese orgány fotosyntézy (listy) a orgány pohlavního rozmnožování (květy, plody). Stonek dále rozvádí roztoky minerálních látek z kořenů do listů a organické asimiláty z listů do růstových pletiv. Neméně důležitá je i skutečnost, že umožňuje růst rostliny, na něm se opět podílejí dělivá pletiva, umístěná ve vzrostném vrcholu. Stonek také pomáhá orientovat listy ke světlu a vyzdvihuje květy nad ostatní rostlinné části.
Někdy mají stonky i další, méně významné funkce. Mladé stonky s chlorenchymem pod pokožkou mají například funkci asimilační. Některé modifikace stonku mají zásobní funkci.
Typy stonku
- Podle tvaru
Dužnaté stonky
- Lodyha (caulis), bylinný stonek, který nese listy,
- Stvol, stonek, který nenese listy
- Stéblo (culmus), dutý stonek trav s kolénky
- Lodyžka, „stonek“ některých mechorostů
Dřevnaté stonky. Dochází k dřevnatění původně dužnatého stonku. Například kmen, hlavní stonek stromů.
- Podle polohy
- přímý
- poléhavý
- plazivý
- vystoupavý
- ovíjivý
- popínavý
Metamorfózy stonku

Podle specializovaných funkcí se často mění tvar a uspořádání stonku. Existují různé metamorfózy:
- oddenek: podzemní ztlustlý stonek, např. přeslička
- šlahoun: nadzemní výběžky např.: jahodník
- oddenková hlíza: podzemní část stonku ztloustne, má funkci zásobní, např. brambor
- stonková hlíza: ztlustlá nadzemní část stonku, např. kedluben
- úponky: k zachytávání se na podkladě, popínavé rostliny, např. réva vinná
- stonkové trny – kolce: ochrana před býložravci, např. trnka
- zdužnatělé stonky: zásobárna vody, např. u kaktusů
- rozšiřování stonku do plochy: souvisí s asimilační funkcí, mívá podobu listů (listnatec)
- brachyblast: např.: modřín (to je modifikace, nemění se funkce)
Růst a výška stonku
Růst stonku se uskutečňuje dělením buněk na vegetačním vrcholu a dále zvětšováním buněk v internodiích. Tloustnutí stonku se děje buď zvětšováním buněk, nebo vytvářením druhotných pletiv, produkovaných kambiem.
Výška stonku je velmi různorodá. Některé druhy vůbec stonek v pravém smyslu nevytváří (např. okřehek). Naopak, nejvyšší stonek má liána rotan rákosovitý (Calamus rotang), a to 200–300 metrů.
Morfologie stonku
Větvení
Způsob větvení stonku má velký vliv na celkový vzhled (habitus) rostliny. Vývojově nejstarším typem větvení stonku je vidličnaté (dichotomické) větvení, při němž se růstový vrchol rozděluje na dvě shodné poloviny. Je charakteristické pro nižší a výtrusné rostliny, vyskytuje se však i u některých vývojově pokročilejších skupin rostlin (palmy, kaktusy aj.).[1] Pro stonky většiny dnešních rostlin je charakteristické větvení postranní, při němž nová větévka vyrůstá z pupenu v paždí listu. Pokud hlavní stonek pokračuje v růstu, jedná se o monopodiální větvení. V případě sympodiálního větvení přebírá růst postranní větévka.
 (c) Avi Aviv Pikiwiki Israel, CC BY 2.5Vidličnatě větvený kmen palmy dumy thébské
(c) Avi Aviv Pikiwiki Israel, CC BY 2.5Vidličnatě větvený kmen palmy dumy thébské Monopodiální kmen olše lepkavé
Monopodiální kmen olše lepkavé Monochaziálně větvená lípa
Monochaziálně větvená lípa Dichaziální větvení kaliny
Dichaziální větvení kaliny
Redukce stonku
Některé rostliny mají vlivem specializované životní formy stonek velmi redukovaný. To je případ některých vodních rostlin vzplývajících na hladině (okřehek, babelka, drobnička), stejně jako některých plně parazitických rostlin (raflézie, saprie aj.).[2]
U některých epifytických orchidejí (např. Polyradicion lindenii, Campylocentrum pachyrrhizum, Harrisella porrecta) je stonek redukovaný na nepatrnou část nutnou k vývoji květních orgánů, zatímco hlavní část rostlinného těla tvoří zelené, asimilující, dužnaté kořeny.[3]
Extrémním případem redukce stonku i ostatních vegetativních částí je čilské jmelí Tristerix aphyllus, rostoucí na některých druzích sloupcovitých kaktusů. Rostlina část života žije pouze uvnitř hostitele a v této fázi nemá ani listy, ani kořeny nebo stonek. Její tělo je tvořeno pouze tenkými parenchymatickými vlákny bez cévních svazků, proplétajícími se mezi buňkami hostitelské rostliny. V určité fázi života vlákna prorostou blízko k povrchu, kde vytvoří pevnou uzlinu, z níž skrz tuhou pokožku hostitele proráží z adventivního pupene výhon květenství.[3]
 Epifytická orchidej Campylocentrum burchellii
Epifytická orchidej Campylocentrum burchellii Květy Tristerix aphyllus na kaktusu
Květy Tristerix aphyllus na kaktusu Poupě saprie Sapria himalayana, vyrůstající přímo z kořene hostitelské rostliny
Poupě saprie Sapria himalayana, vyrůstající přímo z kořene hostitelské rostliny
Anatomie stonku

Primární, druhotně neztloustlý stonek rostlin je tvořen třemi základními pletivy: pokožkou, základním pletivem středního válce a vodivými pletivy. Hlavní hmotu stonku tvoří parenchym, v němž jsou uložena vodivá pletiva. U primárně rostoucích, bylinných stonků se všechna pletiva zakládají prostřednictvím vrcholového meristému. Při druhotném tloustnutí se z druhotného dělivého pletiva zakládá dřevní hmota (xylém) a lýko (floém).
Pokožka a primární kůra
Povrch bylinného stonku tvoří jednovrstevná pokožka (epidermis) krytá voskovou kutikulou, která zamezuje odparu vody a chrání pokožku před patogeny. Pokožka často nese průduchy a chlupy různých typů. U ponořeně rostoucích vodních rostlin většinou kutikula i průduchy chybějí, zatímco pouštní rostliny mívají pokožku ztloustlou, tvořenou několika vrstvami buněk a krytou tlustou kutikulou, někdy i průhlednou korkovou vrstvou (například některé kaktusy a pryšce). Pod pokožkou je primární kůra (cortex). U některých rostlin je tato kůra tvořena v podstatě jen parenchymem, u jiných má složitou stavbu, na níž se podílejí i různé specializované buňky produkující latex, pryskyřici či sliz, případně krystaly šťavelanu vápenatého nebo křemičitá tělíska. Buňky primární kůry často obsahují chloroplasty a podílejí se na fotosyntéze.[4][5][3][6]
Růstový vrchol
Na vrcholu stonku se nachází vrcholový pupen s dělivým pletivem (meristémem), jehož prostřednictvím se stonek prodlužuje a v němž se tvoří základy budoucích listů (listová primordia). U krytosemenných je růstový vrchol krytý tunikou tvořenou jednou nebo několika vrstvami buněk a obklopující jádro vrcholu, tvořené méně pravidelně orientovanými buňkami. U nahosemenných není tunika jasně diferencovaná. Listy se zakládají z okrajů růstového vrcholu dělením buněk tuniky a jádra růstového vrcholu. V paždí listů se tvoří úžlabní pupeny, z nichž vyrůstají buď postranní větévky, anebo květy či květenství.[4][7]
Střední válec a vodivá pletiva
Hlavní hmotu primárního stonku tvoří střední válec (stélé). Je tvořen převážně parenchymem, v němž jsou uloženy cévní svazky. U řady dvouděložných a řidčeji i u jednoděložných bývá vnitřní válec vyplněný dření obklopenou cévními svazky. V pletivu jsou často sklerenchymatická vlákna. Parenchym středního válce může dřevnatět.[4]
Vodivá pletiva stonku lze rozdělit na ta, která vedou vodu a minerální živiny od kořenů k listům (xylém) a ta, která vedou asimiláty od listů na místo spotřeby spolu s minerálními látkami a také rostlinnými hormony (floém). Trubice xylému jsou tvořené odumřelými, tlustostěnnými buňkami (vodivý sklerenchym), zatímco floém drobnějšími, živými buňkami. U druhotně tloustnoucích stonků se mezi oběma typy vodivých pletiv v cévních svazcích nachází jednovrstevná vrstva dělivých meristematických buněk, kambium.[3][7][6] Uspořádání cévních svazků je pro různé skupiny rostlin charakteristické. V primárních stoncích dvouděložných jsou zpravidla uspořádány do kruhu okolo vnitřní dřeně, zatímco u jednoděložných bývají roztroušené v pletivech středního válce.[4]
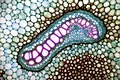 Cévní svazek kapradiny Pteridium pinetorum
Cévní svazek kapradiny Pteridium pinetorum Cévy xylému ve stonku tykve, podélný řez
Cévy xylému ve stonku tykve, podélný řez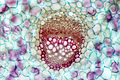 Cévní svazek šťovíku
Cévní svazek šťovíku Roztroušené cévní svazky ve stonku šáchoru Cyperus alternifolius
Roztroušené cévní svazky ve stonku šáchoru Cyperus alternifolius
Druhotné tloustnutí
U všech nahosemenných a u většiny dvouděložných rostlin dochází k druhotnému tloustnutí stonku. Při něm se z vrstvy druhotného meristému, zvaného kambium, zakládají směrem dovnitř stonku vrstvy druhotného xylému, formujícího dřevní hmotu, a směrem vně vrstvy druhotného floému, tvořícího lýko. U dřevin je činnost kambia nejintenzivnější a probíhá po celý život rostliny, zatímco u bylin je dočasná a vytváří jen omezené množství dřevní hmoty. Dřevnatějící stonky jsou kryté druhotnými krycími pletivy, která se vytvářejí dělením buněk felogénu. Jsou tvořena korkovým pletivem, které později vlivem ukládání vrstviček suberinu odumírá a vytváří borku. U jednoděložných rostlin je druhotné tloustnutí výjimečné (juka, dračinec, dračinka) a odlišného charakteru než u dvouděložných a jehličnanů. U palem a pandánů dochází k formování dřevnatého kmene bez účasti sekundárního meristému.[4][8]
 Řez kmínkem tříletého semenáče borovice
Řez kmínkem tříletého semenáče borovice Řez jednoletým stonkem dubu
Řez jednoletým stonkem dubu Řez čtyřletým stonkem bezu
Řez čtyřletým stonkem bezu
Odkazy
Reference
- ↑ GOLA, Edyta M. Dichotomous branching: The plant form and integrity upon the apical meristem bifurcation. Frontiers in Plant Science. June 2014, čís. 5(263). Dostupné online.
- ↑ Obecná botanika: Stavba stonku [online]. Mendelova univerzita v Brně. Dostupné online.
- ↑ a b c d MAUSETH, James D. Botany. An introduction to plant biology. Austin: Jones and Barlett Publishers, 1998. Dostupné online. ISBN 0-7637-0746-5. (anglicky)
- ↑ a b c d e BOWES, Bryan G. A colour atlas of plant structures.. [s.l.]: Manson Publishing, 1999. ISBN 978-1874545163. (anglicky)
- ↑ Obecná botanika: Primární stavba stonku [online]. Mendelova univerzita v Brně. Dostupné online.
- ↑ a b CAPON, Brian. Botany for gardeners. Portland: Timber Press, 2005. Dostupné online. ISBN 0-88192-655-8. (anglicky)
- ↑ a b GLIMN-LACY, Janice; KAUFMAN, Peter B. Botany illustrated. New York: Springer, 2006. Dostupné online. ISBN 0-387-28870-8. (anglicky)
- ↑ SLAVÍKOVÁ, Zdeňka. Morfologie rostlin. Praha: Karolinum, 2002.
Externí odkazy
Média použitá na této stránce
.jpg)
Autor: Berkshire Community College Bioscience Image Library, Licence: CC0
cross section: One year Quercus stem magnification: 100x iron-alum hematoxylin and safranin stain
A cutinized epidermis overlays a two-layer cortex consisting of an outer zone of thick walled collenchyma and an inner zone of loosely packed parenchyma cells. During the first year of growth the underlying activity of the cork cambium begins to replace the epidermis and outer cortical tissues with a protective layer of cork rich periderm. Many cells in the periderm contain dark staining tannins.
Primary growth results in a ring of discrete vascular bundles that divides the parenchymatous cortex and pith. Xylem of each bundle is oriented towards the center of the stem and the phloem to the outside, with fascicular cambium in between. Phloem is heavily capped with sclerenchyma cells and sided by medullary parenchyma.
Due to meristematic activity the vascular bundles expand to form a cylinder consisting of a narrow outer ring of primary phloem, a middle single layered ring of vascular cambium and a deeper large ring of primary xylem. The phloem becomes overlaid by heavy continuous ring of sclerenchyma.
Bulging into the pith are small masses of the first xylem consisting of small protoxylem cells overlaid by a few larger, heavier walled cells of metaxylem. Above this and extending to the cambium are columns of primary xylem divided by narrow rays, extending from the large parenchymatous pith to the phloem.
Autor: , Licence: CC0
Schwarzerle (Alnus glutinosa) an der Saar in Saarbrücken
Autor: RolfDieterMueller, Licence: CC BY 3.0
Hypericum perforatum (Common St John's wort) Stem cross section, stain: Acridine red and Astra blue/Acriflavin.

Autor: Vojtěch Zavadil, Licence: CC BY-SA 4.0
Branch of Viburnum edule, Arboretum Brno, Czech Republic

Autor: Arnoldius, Licence: CC BY-SA 4.0
Winterlinde (Tilia cordata) mit sechs Stämmen im Eingangsbereich des Herner Südfriedhofs an der Wiescherstraße, Naturdenkmal
Autor: based on photo by user Hardyplants ; rewrited, Licence: CC BY-SA 3.0
czech version of "Stem_nodes.jpg" from the English Wikipedia
.jpg)
Autor: Berkshire Community College Bioscience Image Library, Licence: CC0
cross section: Sambucus stem magnification: 100x
During the first year of growth the epidermis is stretched laterally by the expansion of secondary xylem, phloem and cambium. The epidermis is replaced by a protective secondary zone of cork rich periderm. Continual growth of the periderm keeps up with that of underling tissues allowing it to replace the functions of the degrading epidermis.
The outermost layer of periderm consists of layers of cork cells, the phellem, which produce the waterproofing substance suberin. Cork cells are dead at maturity. Deep to the phellem is a layer of living green stained cork cambium or phellogen and just beneath that layers of cork parenchyma or phelloderm. Many cells in the periderm contain dark staining tannins.
In certain areas the cork cambium over produces cork cells, resulting in the formation of ridges and deep cracks in the periderm. These deep fissures, or lenticels, permit gas exchange with tissues under the periderm.
Deep to the periderm is an outer cortex of tightly packed lamellar collenchyma and an inner cortex of open airy parenchyma. Large dark staining tannin ducts are present.
The vascular cylinder consists of a narrow outer of primary and secondary phloem, a middle ring of vascular cambium and a deeper large ring of primary and secondary xylem. Phloem bands of sieve tubes and companion cells are layered and interspaced with parenchyma cells masses and occasional small bundles of sclerenchyma cells. Small masses of calcium oxalate crystals are present. Xylem is separated from the pith by a starch sheath of dark staining parenchyma cells.
The vascular cylinder is divided into narrow columns by radial bands of parenchymatous rays that extend from pith to phloem. Wide phloem rays taper as they dip into the xylem where they merge with the starch sheath. A large parenchymatous pith occupies the center of the stem.
Dark staining tannin producing cells are present throughout the stem including the pith.

.jpg)
Autor: Berkshire Community College Bioscience Image Library, Licence: CC0
long section: Cucurbita stem magnification: 400x
<a href="http://blogs.berkshirecc.edu/bccoer" rel="nofollow">Berkshire Community College Bioscience Image Library</a>
Walls of xylem vessels have lignin free pits and lignin thickenings that form rings, spirals, networks and solid blocks.

Autor: Anatoly Mikhaltsov, Licence: CC BY 4.0
Conductive bunch of rhizomes of fern Pteridium pinetorum ssp. sibiricum. The author used the constant preparation by A.I. Mikhaltsov, polychrome dyeing of tissues according to A.I. Mikhaltsov (2012). As a result, cells of conductive tissues are very clearly visible: large cells whose cell walls are painted in reddish-purple color are cells of xylem, a water-carrying tissue. Cells of the phloem tissue, conducting the products of photosynthesis, are located around the xylem cell. The conductive beam is bounded with cells of the endoderm. Cells of the pericycle are clearly visible between the phloem and endoderm. Parenchyma and sclerenchyma cells (brown cell wall) surround the conductive beam.

Autor: Pato Novoa, Licence: CC BY 2.0
Tristerix aphyllus growing on the columnar cactus, Trichocereus chiloensis
.jpg)
Autor: Len Worthington, Licence: CC BY-SA 2.0
Phu Kieow Wildlife Reserve Chaiyaphum
Thailand.jpg)
Autor: Berkshire Community College Bioscience Image Library, Licence: CC0
cross section: Pinus stem magnification: 40x
During the first year of growth the cutinized epidermis is replaced by protective growth of cork rich periderm. The outer periderm consists of layers of cork cells, the phellem, which produces waterproofing suberin. Cork cells are dead at maturity. Deep to the phellem is a living layer of cork cambium or phellogen and beneath that, layers of cork parenchyma or phelloderm. Many cells in the periderm contain dark staining tannins.
The cortex is divided into a thin outer hypodermis of lignified sclerenchyma cells and thicker inner cortex of thin walled parenchyma cells containing chloroplasts. Large resin ducts are surrounded by secretory parenchyma that produces resins and turpentines. Some cells enlarge into dark staining tyloses.
Both endoderm and pericycle are inconspicuous.
The vascular cylinder or stele in young stems consists of a ring of vascular bundles interspaced with medullary rays of parenchyma cells. Seasonal activity of the cambium replaces the isolated vascular bundles with well-defined annual rings of secondary phloem and xylem. Xylem is endarch with protoxylem found towards center of the stem and younger metaxylem towards the periphery of the stem. Protoxylem consists of annular and spiral tracheids with only tracheids found in metaxylem. True xylem vessels are lacking. Because of the greater production of xylem, the vascular cylinder is dominated by radially arranged rays of secondary xylem interspaced with medullary rays of parenchyma cells. Conspicuous resin ducts are present throughout the xylem.
Phloem is endarch but annual growth the of stem makes it difficult to distinguish between older protophloem to the periphery and younger metapholem towards center of the stem. Phloem lacks companion cells, consisting entirely of sieve tubes and phloem parenchyma. Medullary rays in the secondary phloem include protein rich albuminous cells.
A well-defined pith of parenchyma cells is occasionally interrupted by a few large resin ducts.
Visit the BCC Bioscience Image featuring the Microscopic World of Plants. <a href="http://www.berkshirecc.edu/biologyimages" rel="nofollow">www.berkshirecc.edu/biologyimages</a>
Autor: User:Micropix, Licence: CC BY-SA 3.0
Cyperus alternifolius, stalk, Etzold green.

Autor: Dalton Holland Baptista, Licence: CC BY-SA 3.0
Campylocentrum burchelii